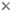Sthenoteuthis oualaniensis
Richard E. Young and Michael Vecchione

Introduction
Sthenoteuthis oualaniensis, commonly known as tobiika, purpleback flying squid or purple squid (among other names), is thought to be the most abundant large squid in the tropical and subtropical waters of the Indo-Pacific region (Young and Hirota, 1998; Dunning, 1998). Its congener, S. pteropus, replaces S. oualaniensis in the Atlantic Ocean.
S. oualaniensis has a complex population structure that consists of three major and two minor forms (Nesis, 1993): A giant form occurs only in the northern Indian Ocean in the region of the Red Sea, Gulf of Aden and Arabian Sea and reaches a maximum size of 650 mm ML; a medium form occurs throughout range of the species and is 120-150 mm ML in mature males and 190-250 mm ML in mature females (this is the "typical" S. oualaniensis); a dwarf form occurs in equatorial waters and lacks the dorsal mantle photophore patch characteristic of the species and is 90-100 mm ML in mature males and 90-120 mm ML in mature females (150 mm ML maximum). The medium form appears to consist of two minor forms based on features of the gladius (double or single lateral axes of the rhachis). One of the two medium forms occurs only in the Red Sea, the Gulf of Aden and the Arabian Sea north of 15°-17°N. Complicating this picture is a small form, similar to the medium form but maturing at a smaller size that is nearly the same size as the dwarf form and is found in the Western Indian Ocean and the eastern tropical Pacific. The dwarf equatorial form is found roughly within 10° latitude of the equator where it co-occurs with the typical S. oualaniensis.
The dwarf form has several morphological characters that separate it from the typical S. oualaniensis: Absence of the dorsal photophore patch, slightly different hectocotylus and slight differences in the spermatophore structure and in the gladius structure (Nesis, 1993). Paralarvae of the dwarf and middle forms cannot be distinguished (Nesis, 1993). Researchers have disagreed on whether or not the dwarf form is a distinct species (Clarke, 1965; Wormuth, 1976; Nesis, 1993). The most recent research on the status of the dwarf form was done by M. Roeleveld-Companyo and she considered it to be a separate species that can only be identified as an adult (personal communication). Snyder (1998) suggests that the giant form results from a plastic phenotype in the species.
Characteristics
- Arms (Wormuth)
- Arm suckers of arm II: Largest suckers with much larger distal teeth than proximal teeth; distal teeth not of uniform size. Distal arm suckers with smooth proximal margins. Basal suckers somewhat irregular. Click on an image to view larger version & data in a new window
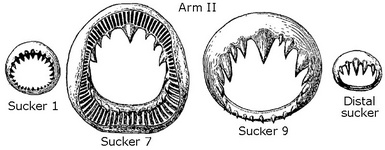
Figure. Oral view of the inner or inner and outer (sucker 7) rings of arm II of S. oualaniensis. Drawings from Sasaki (1929).
Click on an image to view larger version & data in a new window
Figure. Sucker rings from arm II of S. oualaniensis, female, 158 mm ML, Hawaiian waters. Suckers 2 and 9 are tilted to better show proximal edge of ring. Thumbnail (insert) - Shows relative sizes of the three suckers. Photograph by R. Young.
- Hectocotylus (ref)
- Right or left arm IV hectocotylized.
- Hectocotylus with 12-16 suckers.
- Up to six medial pores present on each side (opposite sucker rows 1-6).
- Beyond suckers dorsal membrane decreases then disappears but reappears as thin membrane distally to form groove between it and ventral membrane.
- Ventral membrane similar to dorsal proximally but distally becomes broad and thin.
- Right or left arm IV hectocotylized.
Click on an image to view larger version & data in a new window
Figure. Oral view of the hectocotylus of S. oualaniensis. Arrows point to pores through protective membrane. Photograph by R. Young.
Click on an image to view larger version & data in a new window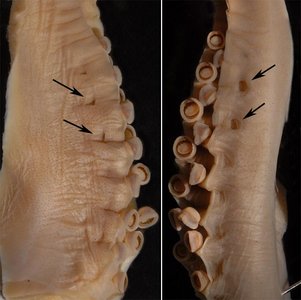
Figure. Hectocotylus of S. oualaniensis, Hawaiian waters. Left - lateral (= dorsal) view. Right - Medial (= ventral) view. Arrows point to pores in protective membrane.
- Arm suckers of arm II: Largest suckers with much larger distal teeth than proximal teeth; distal teeth not of uniform size. Distal arm suckers with smooth proximal margins. Basal suckers somewhat irregular.
- Tentacles
- Carpal locking-apparatus with 3-5 suckers and knobs.
- Usually only one or two toothed suckers found proximal to locking-apparatus.
- Suckers Click on an image to view larger version & data in a new window
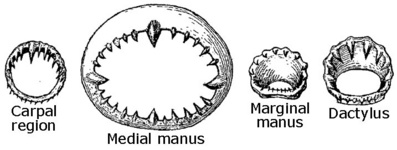
Figure. Oral view of the inner rings of the tentacular club suckers of S. oualaniensis. Drawings from Sasaki (1929).
Click on an image to view larger version & data in a new window

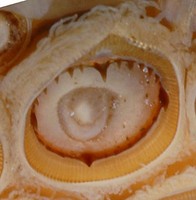
Figure. Oral view of the tentacular club of S. oualaniensis. Top - Entire club. Bottom left - Proximal end of club showing carpal region. Arrows point to carpal pads; carpal suckers located to the immediate right of the pads. Bottom right - Enlarged view of the largest club sucker showing the enlarged tooth in each quadrant. Photographs by R. young.
Click on an image to view larger version & data in a new window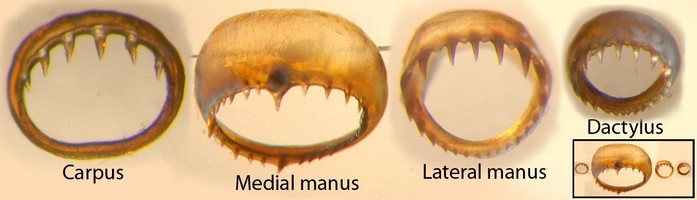
Figure. Sucker rings from the club of S. oualaniensis, female, 159 mm ML, Hawaiian waters. Carpus - Sucker from mid-carpus, opposite second of three locking knobs (this is NOT a locking sucker). Medial manus - One of largest suckers. Lateral manus - Adjacent to largest medial sucker. Dactylus - Mid-dactylus, ventral series. Thumbnail (lower right) - Shows relative sizes of the four suckers. Photograph by R. Young.
- Head
- Beaks: Descriptions can be found here: Lower beak; upper beak.
- Beaks: Descriptions can be found here: Lower beak; upper beak.
- Funnel/mantle locking-apparatus
- Funnel/mantle locking-apparatus fused. A full description of the locking-apparatus can be found here.
Click on an image to view larger version & data in a new window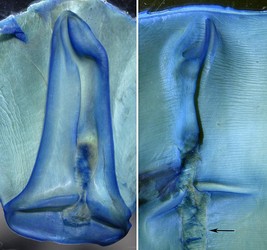

Figure. Frontal view of the funnel component of the funnel/mantle locking-apparatus of S. oualaniensis showing the region where fusion to the dorsal component occurs. Left - Funnel component. Middle - Mantle component. Arrow points to an area where a strip of tissue torn was off the mantle when the two components were broken apart. Right - Anterior tips of the mantle component (top) and funnel component (bottom). Photographs by R. Young.
- Funnel/mantle locking-apparatus fused. A full description of the locking-apparatus can be found here.
- Photophores
- Subcutaneous photophores, including the dorsal-patch, develop at about 100 mm ML. The dwarf form lacks these photophores but has two intestinal photophores.
Click on an image to view larger version & data in a new window
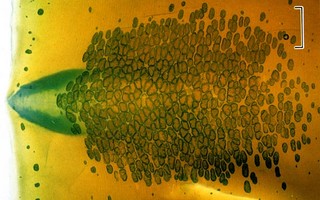
Figure. The mantle of S. oualaniensis that has been cleared and the photophores (and gladius, funnel locking-apparatus) stained with alcian blue stain. The numerous small, dark dots are photophores embedded in the mantle muscle. Left - Ventral (top) and dorsal (bottom) views. Right - Enlargement of the dense patch of several hundred photophores found on the anterodorsal mantle. Photographs (left modified slightly) from Kishimoto and Kohno (1992). Also see photographs on the Sthenoteuthis page.
- Subcutaneous photophores, including the dorsal-patch, develop at about 100 mm ML. The dwarf form lacks these photophores but has two intestinal photophores.
Life History
S. oualaniensis appears to be a "continuous spawner" (Harman, et al., 1989; Young and Hirota, 1998; Roche, et al., 2001) where intermittent spawning occurs over an extended period but accumulation of eggs in the oviducts is a continuous process. A female about 300 mm ML has about 250,000 eggs in the combined, filled oviducts (Harman, et al., 1989). The number of oocytes in various stages of development in a female of 251 mm ML was estimated to be 1,643,000 (Harman, et al., 1989). The length of the spawning period and the frequency of spawning episodes within this period are unknown.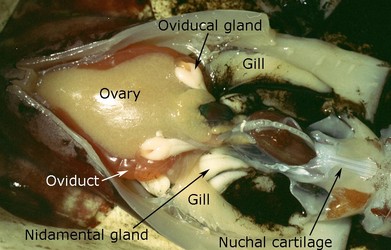
Figure. DORSAL view of the viscera of S. oualaniensis. The large ovary contains developing eggs of many sizes. The oviducts are filled with mature eggs that have an orange tint. The black pigment is ink that has dribbled into the mantle cavity. Photograph by M. Vecchione.
An egg mass, spawned in captivity, by the giant form of S. oualaniensis from the Arabian Sea has a "typical ommastrephid" appearance (Chesalin and Giragosov, 1993) which would be a large spherical, gelatinous egg mass with thousands of eggs distributed thoughout the mass. Mature oocytes (eggs) are small, 0.70 mm x 0.84 mm (Sakurai, et al., 1995).
Paralarvae mostly occupy the upper 50 m of the ocean during both the day and night and often the upper 20 m although some can be found as deep as 80-100 m (Young and Hirota, 1998; Saito and Kubodera, 1993).
In Hawaiian waters paralarvae are present throughout the year but are most abundant in August, less abundant in April and least abundant in October and December (Harman and Young (1985). Some seasonal overlap occurs between the occurrance of paralarvae of S. oualaniensis and the related Ommastrephes bartramii in Hawaiian waters and the Northwestern Pacific; however, most of the spawning of these two species is out of phase in time and space over much of the year (Young and Hirota, 1990; Bower, et al., 1999; Saito and Kubodera, 1993).
Growth of paralarvae based on statolith increments indicates a paralarva of 4 mm ML is 30 days old which is in strong contrast to the growth of O. bartramii which reaches 7 mm ML at 30 days (Bigelow, 1991). Separation of the tentacles, usually thought to mark the end of the paralarval period in ommastrephids, occurred at about 9 mm ML in S. oualaniensis (Harman and Young, 1985). The validity of this event as the marker, however, has been questioned by Yatsu and Mori (2000).
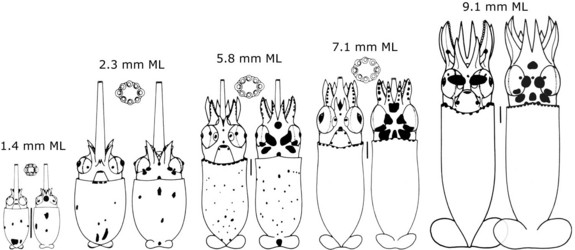
Figure. Ventral and dorsal views of paralarval stages and the first juvenile stage of S. oualaniensis. Chromatophore patterns incomplete. Drawings from Harman and Young (1985).
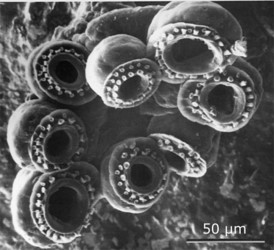
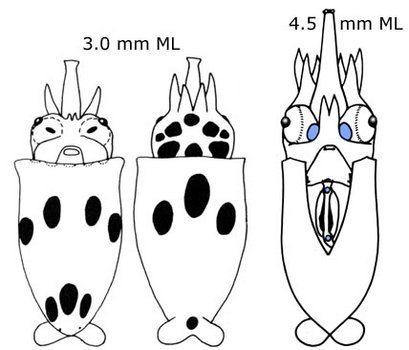
Figure. Paralarval S. oualaniensis. Left - Oral view of the suckers at the tip of the proboscis. Photograph from Harman and Young (1985). Middle - Typical chromatophore pattern at 3.0 mm ML. Drawing from Young and Hirota (1990). Right - Photophore arrangement (blue): ocular photophores and two intestinal photophores. Drawing from Harman and Young (1985).
Juveniles are frequently found aggregated in inshore waters around oceanic islands (Okutani and Tung, 1978). The vertical distribution of juveniles is poorly known. Presumably they occupy near-surface waters during the day and night: Small S. oualaniensis have been found to glide onboard ship during the day (Young and Hirota, 1998), seen gliding over the surface of the ocean during the day (Arata, 1954; Young, 1975; Okutani and Tung, 1978) and found in the stomachs of day feeding birds (Ashmole and Ashmole, 1967). They are also encountered at the surface at night (Young and Hirota, 1998).
The diagnostic photophore patch on the anterodorsal surface of the mantle becomes apparent at approximately 100 mm ML (Kishimoto and Kohno, 1992; Nesis, 1993) and can be taken as a marker for the end of the juvenile phase and the beginning of the subadult phase of the life cycle. The hectocotylus develops about 110 mm ML (Okutani and Tung, 1978). Males of S. oualaniensis reach 15-17 mm in 6-7 months and are thought to live less than a year; the growth rates, determined by growth marks on statoliths and gladii, of the middle and dwarf forms are nearly the same, but the life span is short in the small forms, probably not more than 6 months Nesis (1993).
Yatsu (2000) determined growth curves for both male and female S. oualaniensis based on statolith increments from about 120 mm ML to about 290 mm ML for females and about 185 mm ML for males. His curves are: ML=9.9511X0.634 (females) and ML=29.9X0.3494 (males) with ML in mm and x=increments=days (presumably). According to this curve a female of 120 mm ML is 51 days old. This contrasts strongly with data of Zaidi bin Zakaria (2000), which places a 115 mm ML female at an age of 95 days. Since ommastrephid growth is affected by food availability and ambient temperature, growth rates can vary with season, geographic area and year (Yatsu, 2000).
In Hawaiian waters S. oualaniensis females mature between 158 and 205 mm ML with 50% mature at 166-175 mm ML and 90% mature at 200 mm ML (maximum size is 335 mm ML); males are mostly mature by 140 mm ML (maximum size is 210 mm ML) (Young and Hirota, 1998; Suzuki et al., 1986). In northern Australian waters males reached maturity from 160 mm ML and females from 250 mm ML.
Distribution
Vertical distribution
At night squids are commonly observed in surface waters. suggest, based on the near In Hawaiian waters subadults and adults of S. oualaniensis are usually absent from surface waters at night over bottom depths of less than 650 m,which suggests that they descend to at least 650 m during the day (Young and Hirota, 1998). Simarily, Dunning (1998) caught adult S. oualaniensis in eastern Australian waters only where bottom depths exceeded 600 m.
In many geographical areas of S. oualaniensis distribution, deep water can have low oxygen concentrations which should exclude its presence. Indeed, S. oualaniensis has a very high metabolic rate (standard metabolism of 348 ml O2 kg-1 h-1) that exceeds that of most fast-swimming oceanic fishes (Zuev, et al., 2002). Its energy metabolism, as in other squids, is based mostly on protein; however during metabolism a considerable proportion of the protein is catabolized anaerobically (Shullman et al., 2002). In this sense, these squid may be "preadapted" for low oxygen environments. In the Indian Ocean large S. oualaniensis occupy depths of 300-400 m during the day where the oxygen concentration is 0.1-0.2 mg/liter (2-4% of saturation) (Shullman et al., 2002).
Geographical distribution
S. oualaniensis is a tropical IndoPacific species that occurs in the Pacific from southern Japan to southern Queensland and from just south of Baja California to northern Chile (Nesis, 1987). Dunning (1998) records S. oualaniensis as far south as 38°40'S (SST 20.7°C) at the northern edge of the Bass Straits, near the coast of Australia. In the Coral Sea Basin (east of 155°E), however, adults were caught only north of 32°S (SSTs>23.5°C). Eastward from the Coral Sea to near the coast of South America Wormuth (1976) records its presence roughly along 20°S to about 110°W then his southern records reach about 15°S but stop about 4° of longitude west of Peru apparently at the edge of the Peru current. In the region of the Hawaiian Archipelago, S. oualaniensis did not occur in winter north of 28°N. Between 23°N and 28°N large females were few and of those few most were not mature; the ratio of mature to immature females of squid 200 mm ML or larger was 1:27 during one cruise (Young and Hirota, 1998). This suggests that at the northern end of their range, the females are not maturing normally. Tung et al. (1973) states that Taiwan (which lies roughly between 22 and 25°N) is at "the edge area of the migration sphere of the common squid [S. oualaniensis] in the southwest water of Taiwan... ." S. oualaniensis is recorded off Southern Japan at nearly 35°N (SST 25-26°C) (Saito and Kubodera (1993). Wormuth (1976) recorded the northern limit of distribution at 33.6°N and 165.5°E (September) and about 31°N at 156°W, 139°W and 129°W off Southern California west of the California Current. During the change of seasons the SSTs across most of the Pacific undergo a dramatic north-south movement. For example, north of Hawaii the 18°C isotherm can move from near 3O°N in February to over 40°N in August (Laurs and Lynn, 1991). The extent to which the latitudinal limits of the distribution of S. oualaniensis change with season are not known but do not appear to be very dramatic.
S. oualaniensis exhibits small-scale areal patterns in the vicinity of islands. Around the Hawaiian Islands large mature females are located on the windward (northeastern) sides of the islands, small immature females are located primarily within 25 km of the leeward (southwestern) sides of the islands and somewhat larger mostly immature females are found mainly further offshore on the leeward sides of the islands (Young and Hirota, 1998). The reason for these patterns is unknown but Young and Hirota (1998) found some differences in feeding patterns between windward and leeward areas. These local patterns and those observed in juveniles (Okutani and Tung, 1978) and in paralarvae (Bower, et al., 1999) indicate modified distribution patterns in the vicinity of islands.
References
Ashmole, N. and M. Ashmole. 1967. Comparative feeding ecology of seabirds of a tropical oceanic island. Bull. Peabody Mus. Nat. Hist., 24: 1-131.
Bigelow, K. 1991. Age and growth of three species of squid paralarvae from Hawaiian waters, as determined by statolith microstructures. M.S. Thesis, Univ. of Hawaii, 78 pp.
Bower, J. R. M. P. seki, R. E. Young, K. A. Bigelow, J. Hirota and P. Flament. 1999. Cephalopod paralarvae assemblages in Hawaiian Islands waters. Mar Ecol Prog Ser, 185: 203-212.
Chesalin, M. V. and V. E. Giragosov. 1993. Egg clutch and embryonic development of the squid Sthenoteuthis oualaniensis (giant Arabian form) under experimental conditions. Okeanologiya, 33: 116-120. (In Russian)
Clarke, M. R. 1965. Large light organs on the dorsal surfaces of the squids Ommastrephes pteropus, Symplectoteuthis oualaniensis and Dosidicus gigas. Proc. Malacol. Soc. London, 36: 319-321.
Dunning, M. C. 1998. A review of the systematics, distribution , and biology of the arrow squd genera Sthenoteuthis Verrill, 1880, and Ornithoteuthis Okada, 1927 (Cephalopoda: Ommastrephidae). Smithson. Contr. Zoology, 586: 425-433.
Harman, R. F. and R. E. Young. 1985.The larvae of ommastrephid squids from Hawaiian waters. Vie et Milieu, 35: 211-222.
Harman, R. F., R. E. Young, S. B. Reid, K. M. Mangold, T. Suzuki and R. F. Hixon. 1989. Evidence for multiple spawning in the tropical oceanic squid Sthenoteuthis oualaniensis (Teuthoidea: Ommastrephidae). Mar. Biol., 101: 513-519.
Kishimoto, H. and H. Kohno. 1992. Development of the luminous organ in the purpleback flying squid, Sthenoteuthis oualaniensis, as shown by alcian blue stain techniques. Bull. Inst. Oceanic Res. & Develop., Tokai Univ., 13: 71-83.
Laurs, M. R. and R. J. Lynn. 1991. North Pacific albacore ecology and oceanography. NOAA Tech. Report, No. 105: 69-87.
Nesis, K. N. 1993. Population structure of oceanic ommastrephids, with particular reference to Sthenoteuthis oualaniensis: A review. In: Okutani, T., R. K. O'Dor and T. Kubodera (eds.) Recent Advances in Fisheries Biology. Tokai Univ Press, Tokyo., p. 375-383.
Nesis, K. N. 1982/87. Abridged key to the cephalopod mollusks of the world's ocean. 385+ii pp. Light and Food Industry Publishing House, Moscow. (In Russian.). Translated into English by B. S. Levitov, ed. by L. A. Burgess (1987), Cephalopods of the world. T. F. H. Publications, Neptune City, NJ, 351pp.
Okutani, T. and I. Tung. 1978. Reviews of biology of commercially important squids in Japanese and adjacent waters. I. Symplectoteuthis oualaniensis (Lesson). Veliger, 21: 87-94.
Rocha, F., A. Guerra and A. F. Gonzalez. 2001. A review of reproductive strategies in cephalopods. Biol. Rev., 76: 291-304.
Saito, H. and T. Kubodera. 1993. Distribution of ommastrephid rhynchoteuthion paralarvae (Mollusca: Cephalopoda) in the Kuroshio Region. pp. 457-466. In: Okutani, T., R. K. O'Dor and T. Kubodera (eds.) Recent Advances in Fisheries Biology. Tokai Univ Press, Tokyo.
Sakurai, Y., R. E. Young, J. Hirota, K. Mangold, M. Vecchoine, M. R. Clarke and J. Bower. 1995. Artificial fertilization and development through hatching in the oceanic squids Ommastrephes bartramii and Sthenoteuthis ouananiensis (Cephalopoda: Ommastrephidae). The Veliger, 38: 185-191.
Sasaki, M. 1929. A Monograph of the Dibranchiate Cephalopods of the Japanese and Adjacent Waters. Journal of the College of Agriculture, Hokkaido Imperial University, 20(supplement):357 pages.
Shulman, G., M. Chesalin, G. Abolmasova, T. Yuneva and A. Kidneys. 2002. Metabolic strategy in pelagic squid of genus Sthenoteuthis (Ommastrephidae) as the basis of high abundance and productivity: An overview of the Soviet investigations. Bull. Mar. Sci., 71: 815-836.
Snyder, R. 1998. Aspects of the biology of the giant form of Sthenoteuthis oualaniensis (Cephalopoda: Ommastrephidae) from the Arabian Sea. J. Molluscan Stud., 64: 21-34.
Suzuki, T., S. Yamamoto, K. Ishii and W. Matsumoto. 1986. On the flying squid Stenoteuthis oualaniensis (Lesson) in Hawaiian waters. Bull. Fac. Fish. Hokkaido Univ., 37: 111-123.
Tung. I., C. Lan and C. Hu. 1973. The preliminary investigation for exploitation of common squid resources. Rept. Inst. Fish. Biol. Ministry Economic Affairs and Nat. Taiwan Univ., 3: 211-247.
Wormuth, J. H. 1976. The biogeography and numerical taxonomy of the oegopsid squid family Ommastrephidae in the Pacific Ocean. Bull. Scripps Inst. Oceanogr., 23, 90 pp.
Yatsu, A. 2000. Age estimation of four oceanic squids, Ommastrephes bartramii, Dosidicus gigas, Sthenoteuthis oualaniensis, and Illex argentinus (Cephalopoda, Ommastrephidae) based on statolith microstructure. JARQ-, 34 (1): 75-80.
Yatsu, H. and J. Mori. 2000. Early growth of the autumn cohort of neon flying squid, Ommastrephes bartramii, in the north Pacific Ocean. Fisheries Research, 45: 189-194.
Young, R. E. 1975. A brief review of the biology of the oceanic squid, Symplectoteuthis oualaniensis (Lesson). Comp. Biochem. Physiol. B, 52 (1B): 141-144.
Young, R. E. and J. Hirota. 1990. Description of Ommastrephes bartramii (Cephalopoda: Ommastrephidae) paralarvae with evidence for spawning in Hawaiian waters. Pac. Sci., 44: 71-80.
Young, R. E. and J. Hirota. 1998. Review of the ecology of Sthenoteuthis oualaniensis near the Hawaiian Archipelago. In: Okutani, T. (ed.) Contributed papers to the international symposium on large pelagic squids. Japan Marine Fishery Resources Research Center, Tokyo, p. 132-133.
Zaidi bin Zakaria, M. 2000. Age and growth studies of oceanic squid, Sthenoteuthis oualaniensis using statoliths in the South China Sea, Area 3, Western Philippines. Proc. Third Tech. Sem. Mar. Fish. Res. Survey in the South China Sea, Area 3: Western Philippines, No. 41: 118-134.
Zuev, G., C. Nigmatullin, M. Chesalin and K. Nesis. 2002. Main results of long-term worldwide studies on tropical nektonic oceanic squids genus Sthenoteuthis: An overview of the Soviet investigations. Bull. Mar. Sci., 71: 1019-1060.
Title Illustrations

| Scientific Name | Sthenoteuthis oualaniensis |
|---|---|
| Location | Hawaiian waters |
| Specimen Condition | Live Specimen |
| View | Side |
| Size | Estimate of 18-20 cm ML |
| Image Use |
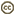 This media file is licensed under the Creative Commons Attribution-NonCommercial License - Version 3.0. This media file is licensed under the Creative Commons Attribution-NonCommercial License - Version 3.0.
|
| Copyright |
© 1996

|
About This Page
University of Hawaii, Honolulu, HI, USA
National Museum of Natural History, Washington, D. C. , USA
Page copyright © 2015 and
 Page: Tree of Life
Sthenoteuthis oualaniensis .
Authored by
Richard E. Young and Michael Vecchione.
The TEXT of this page is licensed under the
Creative Commons Attribution-NonCommercial License - Version 3.0. Note that images and other media
featured on this page are each governed by their own license, and they may or may not be available
for reuse. Click on an image or a media link to access the media data window, which provides the
relevant licensing information. For the general terms and conditions of ToL material reuse and
redistribution, please see the Tree of Life Copyright
Policies.
Page: Tree of Life
Sthenoteuthis oualaniensis .
Authored by
Richard E. Young and Michael Vecchione.
The TEXT of this page is licensed under the
Creative Commons Attribution-NonCommercial License - Version 3.0. Note that images and other media
featured on this page are each governed by their own license, and they may or may not be available
for reuse. Click on an image or a media link to access the media data window, which provides the
relevant licensing information. For the general terms and conditions of ToL material reuse and
redistribution, please see the Tree of Life Copyright
Policies.
- First online 29 November 2009
- Content changed 10 November 2012
Citing this page:
Young, Richard E. and Michael Vecchione. 2012. Sthenoteuthis oualaniensis . Version 10 November 2012 (under construction). http://tolweb.org/Sthenoteuthis_oualaniensis/77447/2012.11.10 in The Tree of Life Web Project, http://tolweb.org/