



Baphetidae
Loxommatidae
Jennifer A. Clack
- Baphetes bohemicus

- Baphetes kirkbyi
- Baphetes lintonensis
- Baphetes planiceps
- Eucritta melanolimnetes
- Kyrinion martilli
- Loxomma acutirhinus
- Loxomma allmani
- Loxomma rankini
- Megalocephalus lineolatus
- Megalocephalus pachycephalus
- Spathicephalus mirus
- Spathicephalus pereger
Introduction
Among the most bizarre and enigmatic tetrapods of the Carboniferous period are the baphetids (formerly known as loxommatids, see Milner and Lindsay (1998)). Though their skulls are not uncommon, there is little yet known about their postcrania. Baphetids were among the first of the Carboniferous fossil tetrapods to be found and were originally described in 1850 by William Dawson. They ranged from the Late Mississippian to the Middle Pennsylvanian, a period of about 35 million years. Most specimens come from Scotland and northern England (Beaumont 1977).

Figure 1. Skull of Kyrinion martilli, a recently discovered baphetid from the north east coast of England. Hancock Museum, Newcastle upon Tyne, UK. Copyright © 1997 Hancock Museum.
Discussion of Phylogenetic Relationships
Baphetids were at one time regarded as relatives of temnospondyls, but since the advent of cladistic analysis, it has been suggested that they show no derived characters in common with this group (eg. Panchen 1980). They have been placed as relatives of the reptiliomorph group (amniotes and extinct related groups) as defined by Panchen and Smithson (1988), but the characters on which this grouping was founded are seen more widely among stem-tetrapods. When data from Eucritta is included, baphetids appear as either sister to temnospondyls or to anthracosaur, more or less with equal frequency (see Eucritta melanolimnetes). Some characters of the ear and occiput have recently been described that add further support to the original view of a relationship with temnospondyls (Clack 2003). Work in progress by Robinson one the otic region and stapes is also likely to support this suggestion. Ruta et al. (2003) placed baphetids as a stem tetrapod plesion falling immediately below temnospondyls. No analysis of the interrelationships of the constituent taxa has ever been formally proposed, though work by Milner and Walsh (in preparation) should resolve some of the issues.
The author disagrees with the move to restrict the vernacular term ‘tetrapod’ to a crown clade (Gauthier et al., 1989). In this page, the term ‘tetrapod’ and ‘stem-tetrapod’ refer only to vertebrates with limbs and digits. (Refer to the page on the Definition of the taxon Tetrapoda to get more information on this topic.)
Characteristics
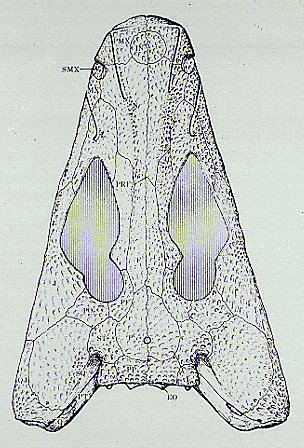
There are six baphetid genera but very few unique characters unite them, while they share many primitive characters such as a closed palate with fang-pairs on the marginal palatal bones. All the baphetids had a strongly embayed spiracular notch, while the stapes was distally broad (Beaumont and Smithson 1998, Clack 2003). One distinctive feature they share is a characteristic ‘key-hole shaped orbit’, formed by excavation of the lacrimal and prefrontal anterior to the orbit (see title figure). Few other characters unite the four genera apart from those found more widely in stem-tetrapods. In the basal baphetid Eucritta, this feature is incipiently shown (Clack 1998, 2001)
The baphetid orbit
The function of the anterior extension of the orbit in baphetids has stimulated much speculation. One suggestion is that the space was occupied by a salt-gland, ridding the body of excess salt. The bone around the rim however, is excavated on the internal suface, which suggests that any gland was housed inside the skull. The coal swamps in which baphetids lived are usually considered to have been freshwater, though recent work (Schultze et al. 1994) has suggested that some of them were brackish, so that presence of a salt gland might be not be considered so unlikely.
Another possibility is that the space was occupied by the pterygoideus muscle used for rapid closing of the lower jaw . With the shallow skull characteristic of baphetids, there may have been little room for this muscle, which is thought to have run dorsally along the inside of the palate, to bulge during contraction. Extra room may have allowed the muscle to have developed more power for snapping the jaws shut. Alternatively, the muscles may have originated around the rim of the hole, giving a firmer attachment. However, the course of the muscle is conjectural (Beaumont 1977).
A third possibility is that the space was occupied by an electrosensory organ (Bjerring, 1986). Such organs are not uncommon in modern fishes which live in murky vegetation-choked water, and they help the animals to detect electrical impulses produced by the muscles of fishes or tetrapods, when vision is of limited use.
References
Beaumont, E. I. (1977) Cranial morphology of the Loxommatidae (Amphibia: Labyrinthodontia). Philosophical Transactions of the Royal Society, London, B, 280: 29-101.
Beaumont, E. I., and Smithson, T. R. (1998) The cranial morphology and relationships of the aberrant Carboniferous amphibian Spathicephalus mirus Watson. Zoological Journal of the Linnean Society of London.122: 187-209.
Bjerring H. C. 1986. Electric tetrapods? In: Z. Rocek (ed.) Studies in Herpetology. Prague, Charles University, pp. 29-36.
Clack, J. A. 1998 A new Early Carboniferous tetrapod with a mélange of crown group characters. Nature 394: 66-69
Clack, J. A. 2001 Eucritta melanolimnetes from the Early Carboniferous of Scotland, a stem tetrapod showing a mosaic of characteristics. Trans R. Soc. Ed. 92, 75-95
Clack, J. A. 2003 A new baphetid (stem tetrapod) from the Upper Carboniferous of Tyne and Wear, UK, and the evolution of the tetrapod occiput. Can. J. Earth Sci. 40: 483-498
Gauthier, J., Canatella, D., De Queiroz, K., Kluge, A. and Rowe, T. (1989). Tetrapod phylogeny. In: Fernholm, B., Bremer, K. and Jornwall, H. (Eds), The Hierarchy of Life. London, Elsevier Science Publishers, pp. 337-353.
Milner, A. C. and Lindsay, W (1998) Postcranial remains of Baphetes and their bearing on the relationships of the Baphetidae (=Loxommatidae). Zoological Journal of the Linnean Society of London. 122: 211-235.
Panchen, A. L. 1980 The origin and relationships of the anthracosaur Amphibia from the Late Palaeozoic. In Panchen, A. L. (ed) The terrestrial environment and the origin of land vertebrates. Sys. Ass. Spec. Vol. 15. Academic Press, London, pp. 319-350.
Panchen A. L., Smithson T. R. 1988. The relationships of the earliest tetrapods. In M. J. Benton. ed. The phylogeny and classification of the tetrapods. Oxford: Clarendon Press, pp. 1-32.
Ruta, M., Coates, M. I. & Quicke, D. L. J. 2003. Early tetrapod relationships revisited. Biological Reviews 78: 251-345.
Schultze, H-P., Maples, C. G. and Cunningham, C. R. 1994. The Hamilton Konservat-Lagerstatte: Stephanian terrestrial biotas in a marginal-marine setting. Transactions of the Royal Society of Edinburgh, Earth Sciences, 84: 249-260.
Title Illustrations

| Scientific Name | Megalocephalus |
|---|---|
| Comments | skull reconstruction |
| Reference | Beaumont, E. I. (1977) Cranial morphology of the Loxommatidae (Amphibia: Labyrinthodontia). Philosophical Transactions of the Royal Society, London, B, 280: 29-101. |
| Acknowledgements | reproduced with permission |
| Copyright | © 1977 Royal Society, London |
About This Page
I would like to express my grateful thanks to Michel Laurin for guiding me through the production of these pages and for editing the result. I would also like to thank David Maddison for scanning in the images for me, and Katja Schulz for finalising the appearance of the pages.
Jennifer A. Clack

University Museum of Zoology, Cambridge, UK
Correspondence regarding this page should be directed to Jennifer A. Clack at
Page copyright © 2011 Jennifer A. Clack
 Page: Tree of Life
Baphetidae. Loxommatidae.
Authored by
Jennifer A. Clack.
The TEXT of this page is licensed under the
Creative Commons Attribution-NonCommercial License - Version 3.0. Note that images and other media
featured on this page are each governed by their own license, and they may or may not be available
for reuse. Click on an image or a media link to access the media data window, which provides the
relevant licensing information. For the general terms and conditions of ToL material reuse and
redistribution, please see the Tree of Life Copyright
Policies.
Page: Tree of Life
Baphetidae. Loxommatidae.
Authored by
Jennifer A. Clack.
The TEXT of this page is licensed under the
Creative Commons Attribution-NonCommercial License - Version 3.0. Note that images and other media
featured on this page are each governed by their own license, and they may or may not be available
for reuse. Click on an image or a media link to access the media data window, which provides the
relevant licensing information. For the general terms and conditions of ToL material reuse and
redistribution, please see the Tree of Life Copyright
Policies.
- First online 28 January 1998
- Content changed 09 February 2006
Citing this page:
Clack, Jennifer A. 2006. Baphetidae. Loxommatidae. Version 09 February 2006. http://tolweb.org/Baphetidae/15011/2006.02.09 in The Tree of Life Web Project, http://tolweb.org/