



Ichthyostega
Jennifer A. Clack

Probable synonyms: I. watsoni I. eigili I. kochi Ichthyostegopsis wimani
Introduction
Ichthyostega specimens were the first Devonian tetrapods to be found and described (Säve-Söderbergh 1932, Jarvik 1952, 1980, 1996). Ichthyostega comes from the Upper Devonian (Famennian) of East Greenland and is widely featured in the literature as the first ‘four-legged fish’. Reconstructions of the animal (see title figure) have been influential in ideas about what the earliest tetrapods were like, though more recent studies (Bjerring 1985, Coates and Clack 1990, Clack et al. 2003, Ahlberg et al. 2005) have challenged more traditional interpretations (Jarvik 1996).

Figure 1. Skull roof of Ichthyostega. MGUH VP 6064 Geological Museum, Copenhagen. Photo J. A. Clack. Copyright © 1997 University Museum of Zoology, Cambridge.
Phylogenetic Position of Ichthyostega
Four species were originally named by Save-Soderbergh (1932), and a second genus, Ichthyostegopsis was also described. Jarvik (1996) suggested that these were most likely to be synonyms but further study has shown the existence of three valid species based on skull proportions and associated with three distinct horizons in the Celsius Bjerg Formation: I. stensioei, I. eigili, and I. watsoni (Blom 2005). Ichthyostegopsis is probably a juvenile I. eigili. Ichthyostega shows a mixture of plesiomorphic and apomorphic characters which make it difficult to place phylogenetically, though it is clearly a basal stem-tetrapod (Ahlberg and Clack 1998, Clack 2002a, Ruta et al. 2003). Only one character, lack of an intertemporal (Fig. 2), may indicate a close relationship to Acanthostega, but the phylogenetic siginifcance of this feature is debatable. There is certainly no basis for the clade 'Ichthyostegalia'.
The author disagrees with the move to restrict the vernacular term ‘tetrapod’ to a crown clade (Gauthier et al. 1989). In this page, the term ‘tetrapod’ and ‘stem-tetrapod’ refer only to vertebrates with limbs and digits. (Refer to the note on the Definition of the taxon Tetrapoda to get more information on this topic.)
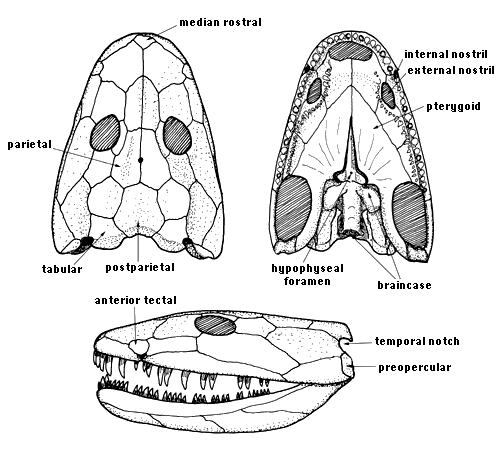
Figure 2. Reconstructions of the skull of Ichthyostega in dorsal (top left), ventral (top right) and lateral views (bottom). © 1997 J. A. Clack.
Skull
The skull roof was broad and flat with a rounded snout. The structure retains some primitive characters such as a median rostral, and an anterior tectal above the external nostril, but recent studies have been unable to confirm the presence a lateral rostral that carried the lateral line canal beneath the nostril (cf. Jarvik 1980). The skull table was broad, lacking an intertemporal but with a pentagonal supratemporal, unlike the dart-shaped bone of Acanthostega. There was a single midline postparietal. The tabulars, also approximately pentagonal, each bore a posteroventral notch and had deep ventrally projecting flanges which attached to the braincase.
The closed palate bore a row of teeth of varying sizes in the palatal arcade, but though some are relatively large, the distinct fang pairs found in many other stem-tetrapods are not always distinguishable or consistently present.
The braincase does not resemble those of other tetrapods or osteolepiforms, such that it has only recently proved possible to identify the anatomical regions with any confidence. The otic region was elongated, with a narrow, notochordal basioccipital with no occipital condyle. The braincase and ear region were very highly modified, with enclosed apparently air-filled chambers either side of the otic region. The delicate, spoon-shaped stapes projected into the chambers, cupping the its floor. It articulated with the basicoccipital via a narrow stem and footplate that occluded a small foramen, presumably the homologue of the fenestra vestibuli. The ear apparatus has been interpreted as adapted for underwater audition (Clack et al. 2003).
Ichthyostega is now known to have had an ossified hyobranchial skeleton comparable to that in Acanthostega (Clack et al. 2003), but recent work has not confirmed the presence of a subopercular, rather this bone may have been a misidentified element of the hyobranchial skeleton.
The lower jaw consists of the usual complement of dermal bones, with three coronoids bearing a row of small thorn-shaped teeth, a parasymphysial plate, and a long prearticular that bore no shagreen. There was a disparity between the upper and lower dentitions, with the lower jaw having about 37 toothspaces, against about 32 larger teeth in the upper jaw.
Postcranial skeleton
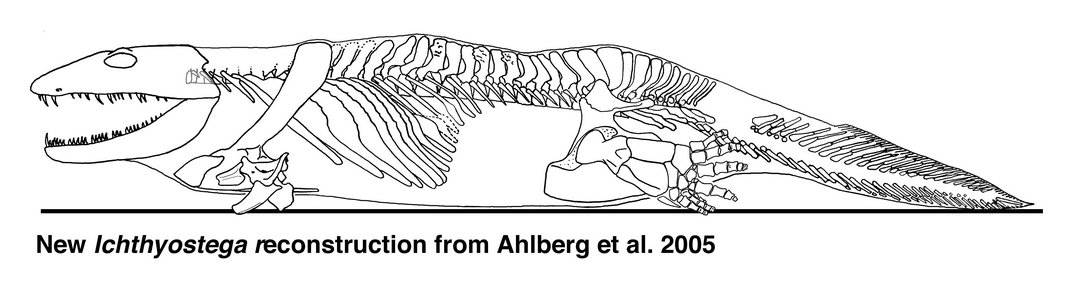
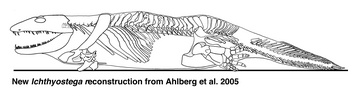
The most conspicuous feature of the axial skeleton is the degree to which the ribs overlapped each other. One thoracic rib may overlap three or four more posterior ones, forming a barrel-shaped ‘corset’ around the body. This is in contrast to the short straight ribs of Acanthostega. It suggests that the animal could not flex its trunk laterally during either walking or swimming. The vertebrae were notochordal, but the neural arches bore more prominent zygapophyses than Acanthostega. Recent work has shown that there was considerable differentiation among the neural arches, particulary of the presacral and sacral regions. The attitude of the zygapophyses in these regions, combined with overlapping ribs has suggested that the animal moved more by dorsoventral flexion than by the conventional lateral walk. The massive forelimbs may have been used to pull the animal forwards followed by flexion of the presacral region to pull up the rear, in what has been described as an 'inch-worm' like mode of locomotion (Ahlberg et al. 2005). Their reconstruction, shown above, is very different from earlier versions (cf. Jarvik 1980, 1996).
The atlas and axis arches are unknown. The tail, shorter than shown by Jarvik (1996), bore a fin supported by supraneural spines but fewer, shorter fin-rays than in Acanthostega.
In the shoulder girdle, the cleithrum and clavicle were co-ossified as in Acanthostega and Hynerpeton. There is no evidence for a post-branchial lamina or anocleithrum. Large supraglenoid and supracoracoid foramina pierced the scapulocoracoid.
The humerus was L-shaped and flattened, but had a more prominent ectepicondylar buttress than in most other tetrapods. The bone was pierced by several accessory foramina lost in most other tetrapods. The radial condyle was long and positioned anteroventrally, while the ulna condyle was terminal. The ulna bore a conspicuous, forked olecranon process, while the radius was short and stubby. In these forearm bones, Ichthyostega resembles later stem-tetrapods more closely than Acanthostega. The wrist and manus are unknown.
The pelvic girdle was large and unitary in structure. It bore a horizontal post-iliac process and a dorsally orientated iliac blade. The sacral rib is undescribed.

Figure 3. Hindlimb of Ichthyostega; specimen collected in 1987. Geological Museum, Copenhagen MGUH Field number 1349. Photo M. Coates. Copyright © 1997 University Museum of Zoology, Cambridge.
The hindlimb consisted of a short, stout femur with a large adductor flange and deep intercondylar fossa (Figs 3 and 4). The large, almost quadrangular tibia and shorter fibula were both flattened. A large intermedium and fibulare comprised most of the identifiable ankle bones; tarsals and metatarsals are difficult to recognize. A well-preserved specimen collected in 1987 shows a full complement of seven digits, three small ones at the leading edge and four stout ones more posteriorly (Coates and Clack 1990). Jarvik (1996) gave a different interpretation. The new interpretation is more consistent with recent discoveries about the axial skeleton.
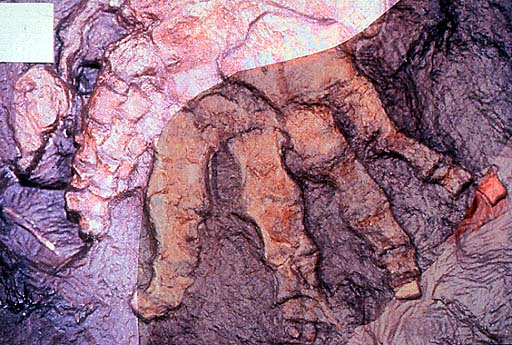
Figure 4. Photographs of part and counterpart superimposed to show seven digits (specimen collected in 1987). Geological Museum, Copenhagen MGUH Field number 1349. Photo M. Coates. Copyright © 1997 University Museum of Zoology, Cambridge.
Paleoecology and lifestyle
No Ichthyostega specimen has been described in its sedimentological context, but the material discovered in 1987 appears to have derived from a channel deposit. Specimens are frequently found in association with Remigolepis plates and Holoptychius scales, but the ecological association between these forms is not understood (Clack and Neininger 2000, Blom et al. 2005). Many Ichthyostega specimens are found in coarse red matrix, while the best-preserved come from a much finer-grained, darker red rock. All appear to have been transported to some degree, and are preserved in flood deposits. Ichthyostega and Acanthostega specimens are not, as a rule, found in quite the same horizons, but the precise temporal and ecological separation between them has yet to be worked out.
In Ichthyostega, the forelimbs seem to have been weight-bearing, but the forearm was unable to extend fully. The proportions of an elephant seal appear the closest analogue among living animals. Perhaps Ichthyostega hauled itself up shelving beaches, moving its forelimbs in parallel and dragging its hindquarters along (Clack 2002b, Ahlberg et al. 2005).
Other Names for Ichthyostega
- Ichthyostega stensioei
- Ichthyostega eigili
- Ichthyostega watsoni
- Ichthyostegopsis wimani
References
Ahlberg, P. E. and Clack, J. A. 1998 Lower jaws, lower tetrapods: a review based on the Devonian tetrapod Acanthostega. Trans. Roy. Soc. Edinb. 89: 11-46
Ahlberg, P. E., Clack, J. A. and Blom, H. 2005 The axial skeleton of the Devonian tetrapod Ichthyostega. Nature. 437, 137-140.
Blom, H., 2005. Taxonomic revision of the Late Devonian tetrapod Ichthyostega from Greenland. Palaeontology. 48, 111-134.
Blom, H., Clack, J. A. & Ahlberg, P. E. 2005. Localities, distribution and stratigraphical context of the Late Devonian tetrapods of East Greenland. Meddelelser om Grønland, Geosciences. 43, 1-50.9
Bjerring, H. C. 1985. Facts and thoughts on piscine phylogeny. In Evolutionary Biology of Primitive Fishes. Edited by R. E. Foreman, A. Gorbman, D. J. M. and R. Olsson, pp. 31-57. Plenum Publishsing Corporation.
Clack, J. A. 2002a An early tetrapod from Romers Gap. Nature 418, 72-76.
Clack, J. A.2002b Gaining ground: the origin and evolution of tetrapods. Indiana University Press, Bloomington, Indiana, USA. 369pp
Clack, J. A., Ahlberg P. E., Finney, S. M, Dominguez Alonso P., Robinson, J and Ketcham, R. A. 2003. A uniquely specialised ear in a very early tetrapod. Nature. 425, 65-6
Clack, J. A and Neininger, S. L. 2000 Fossils from the Celsius Bjerg Group, Upper Devonian sequence, East Greenland: significance and sedimentological distribution. P. F. Friend and B. Williams (Eds) New perspectives on the Old Red Sandstone. Geological Society, London Special Publication 180, 557-566.
Coates, M. I., and J. A. Clack. 1990. Polydactyly in the earliest known tetrapod limbs. Nature, 347:66-69.
Gauthier, J., D. Canatella, K. De Queiroz, A. Kluge, and T. Rowe. 1989. Tetrapod phylogeny. In The Hierarchy of Life. Edited by B. Fernholm, K. Bremer and H. Jornwall, pp. 337-
353. London: Elsevier Science Publishers.
Jarvik, E. 1952. On the fish-like tail in the ichthyostegid stegocephalians. Meddelelser om Grønland, 114(12):1-90.
Jarvik, E. 1980. Basic Structure and Evolution of Vertebrates. Volumes 1 and 2. New York: Academic Press.
Jarvik, E. 1996. The Devonian tetrapod Ichthyostega. Fossils and Strata, 40: 1-213.
Ruta, M., Coates, M. I. & Quicke, D. L. J. 2003. Early tetrapod relationships revisited. Biological Reviews 78: 251-345
Säve-Söderbergh, G. 1932. Preliminary note on Devonian stegocephalians from East Greenland. Meddelelser om Grønland, 98(3):1-211.
Title Illustrations
| Scientific Name | Ichthyostega |
|---|---|
| Comments | Skeletal reconstruction of Ichthyostega based on recent finds and a new interpretation of the hind limb anatomy and proportions. |
| Specimen Condition | Fossil -- Period: Devonian |
| Image Use |
 This media file is licensed under the Creative Commons Attribution-NonCommercial License - Version 3.0. This media file is licensed under the Creative Commons Attribution-NonCommercial License - Version 3.0.
|
| Copyright |
© 2005 Jennifer A. Clack

|
About This Page
I would like to express my grateful thanks to Michel Laurin for guiding me through the production of these pages and for editing the result. I would also like to thank David Maddison for scanning in the images for me, and Katja Schulz for finalising the appearance of the pages.
Jennifer A. Clack
University Museum of Zoology, Cambridge, UK
Correspondence regarding this page should be directed to Jennifer A. Clack at
Page copyright © 2011 Jennifer A. Clack
 Page: Tree of Life
Ichthyostega.
Authored by
Jennifer A. Clack.
The TEXT of this page is licensed under the
Creative Commons Attribution-NonCommercial License - Version 3.0. Note that images and other media
featured on this page are each governed by their own license, and they may or may not be available
for reuse. Click on an image or a media link to access the media data window, which provides the
relevant licensing information. For the general terms and conditions of ToL material reuse and
redistribution, please see the Tree of Life Copyright
Policies.
Page: Tree of Life
Ichthyostega.
Authored by
Jennifer A. Clack.
The TEXT of this page is licensed under the
Creative Commons Attribution-NonCommercial License - Version 3.0. Note that images and other media
featured on this page are each governed by their own license, and they may or may not be available
for reuse. Click on an image or a media link to access the media data window, which provides the
relevant licensing information. For the general terms and conditions of ToL material reuse and
redistribution, please see the Tree of Life Copyright
Policies.
- First online 28 January 1998
- Content changed 09 February 2006
Citing this page:
Clack, Jennifer A. 2006. Ichthyostega. Version 09 February 2006. http://tolweb.org/Ichthyostega/15015/2006.02.09 in The Tree of Life Web Project, http://tolweb.org/