Phyllotini
Leaf-eared mice and their relatives
Scott J. Steppan




This tree diagram shows the relationships between several groups of organisms.
The root of the current tree connects the organisms featured in this tree to their containing group and the rest of the Tree of Life. The basal branching point in the tree represents the ancestor of the other groups in the tree. This ancestor diversified over time into several descendent subgroups, which are represented as internal nodes and terminal taxa to the right.

You can click on the root to travel down the Tree of Life all the way to the root of all Life, and you can click on the names of descendent subgroups to travel up the Tree of Life all the way to individual species.
For more information on ToL tree formatting, please see Interpreting the Tree or Classification. To learn more about phylogenetic trees, please visit our Phylogenetic Biology pages.
close boxTree based on Steppan (1993; 1995; unpubl. ms.; unpubl. data), Steppan and Pardi?as (unpubl. ms.), and Braun and Mares (1995).
Introduction
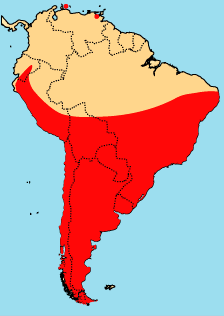
The phyllotines are among the most diverse group of New World muroids (mice and rats) in terms of external appearance and habitats. They range in size from about 12 grams (Calomys hummelincki) to 160 grams (Chinchillula), have tails from 35% (Galenomys) to 145% (Irenomys) of body length, fur color that varies light sandy with pure white undersides in one desert species (Phyllotis gerbillus) to nearly uniform dark brown in a forest species (Loxodontomys). As just indicated, members live in a range of habitats, but are concentrated in open, pastoral habitats and do not occur in tropical or montain rainforests. They reach maximum diversity in the Andean altiplano, where approximately 44% of all phyllotine species can be found (Reig, 1986). Nearly the entire elevational range is encompased by a single species, Phyllotis xanthopygus, from sea level to over 5000 m (16,400'). Phyllotis xanthopygus is possibly the most widespread species, but most other species have much more restrictred distributions, perhaps due to habitat specializations. The combination of being relatively accessible for study and the high local abundances of many species has resulted in the phyllotines being better understood than any other sigmodontine group.
Characteristics
The phyllotines generally share simplified molars for a diet emphasizing plants and seeds, while the skull and other features are relatively generalized. Although most authors have considered the phyllotines to be a relatively well-defined and "cohesive" group, there are actually few convincing shared derived features that define the tribe. These putative synapomorphies include complete loss of the mesoloph and mesolophid (a transformation occuring repeatedly among sigmodontines), premaxillary subequal in posterior extent with nasals, large sphenopalatine vacuity, mesopterygoid narrower than parapterygoid, and large pinnae (ears). Other possible snyapomorphies include the zygomatic arch inserting high on the rostrum and moderately long interparietal. Other characteristic features that are probably plesiomorphic are a hairy heel, a gall bladder, long palate, long incisive foramina, angled or ridged suprorbital region, deeply incised zygomatic notch, and moderate to large third molars. Most species (except Calomys sensu stricto) share the derived second pair of preputial glands (found in the prepuce of the penis) which apparently evolved early within the tribe's diversification.
Discussion of Phylogenetic Relationships
An alternate hypothesis was proposed by Braun (1993).
======================================= Pseudoryzomys
|
| ==================================== Calomys
| |
| | ================================= Eligmodontia
| | |
| | | |||||||||||||||||||||||||||||| Paralomys (Phyllotis amicus and gerbillus)
| | | |
| | | | |||||||||||||||||||||||| Graomys
| | | | ===|
| | | | | ======================== Andalgalomys
| | | | |
| | | | | |||||||||||||||||||||||| Phyllotis
| | | | | |
| | | | | | ===================== Irenomys
| | | | | | |
=====| | | | | | | ================== Euneomys
===| | | | | | |
===| | | | | | |||||||||||| Auliscomys
===| | | | | ===|
===| | | | | ============ Galenomys
| | | | |
===| | | | ============ Loxodontomys
===| | | |
===| | | ====== Neotomys
===| | ===|
| | | ====== Reithrodon
===| |
===| ====== Punomys
| |
===| === Andinomys
===|
=== Chinchillula
Figure 1. Braun's hypothesis is a composite of parsimony analyses conducted on taxonomic subsets so as to maximally resolve the tree. Because Phyllotis amicus and gerbillus were so basal, she resurrected Paralomys to encompass them. To accomodate a paraphyletic Auliscomys, she created Maresomys for A. boliviensis, leaving the remaining two species to form a monophyletic Auliscomys. Loxodontomys was resurrected to contain (Auliscomys) micropus. Steppan (1993) also presented a morphology-based phylogenetic hypothesis, although including a broader survey of characters. A subsequent reanalysis (Steppan, 1995b) resulted in the following hypothesis.
||||||||||||||||||||||||||||||| Calomys
|
| |||||||||||||||||||||| Phyllotis
| |
| | ||| Graomys
| ======| ===|
| | ==Graomys group=| === Andalgalomys
| | |
| | ====== Eligmodontia
| |
| | ====================== Auliscomys
| |=====|
=====| | ====================== Galenomys
===|
|=========================== Chinchillula
|
|=========================== P. wolffsohni
|
| ====================== Irenomys
|=====|
| ====================== Andinomys
|
| ====================== Loxodontomys
======|
====================== Reithrodon group
Figure 2. This tree represents an 80% majority rule tree wherein Punomys is not a phyllotine. In the most-parsimonious tree, which is one step shorter, Punomys is placed as the sister group to Irenomys/Andinomys, while not changing any of the other relationships. This tree was favored because a phylogenetic analysis of the family sigmodontinae indicated that Punomys was not a phyllotine. That same sigmodontine analysis definitively excluded Pseudoryzomys from the phyllotines and placed it in Oryzomyini. Calomys sorellus was most likely more closely related to the remaining phyllotines, but most interior nodes were not well supported. Phyllotis was polyphyletic and had the Graomys group arising from within in, most closely related to P. amicus and gerbillus. Basal nodes within Phyllotis were so weakly supported, however, that no taxonomic changes were recommended. In fact, trees only a few steps longer placed the Graomys group between Calomys and the remaining phyllotines in much the configuration show at the top of this page. Paraphyly of Graomys with respect to Andalgalomys resulted in synonomy of the latter.
Generic Portraits
Compare the skuls, jaws, or molars of all phyllotine genera by selecting the appropriate view below.
References
Baskin, J. A. 1978. Bensonomys, Calomys, and the origin of the phyllotine group of neotropical cricetines (Rodentia: Cricetidae). J. Mamm. 59:125-135.
Baskin, J. A. 1986. The late Miocene radiation of Neotropical sigmodontine rodents in North America. 3:287-303.
Braun, J. K. 1993. Systematic relationships of the tribe Phyllotini (Muridae: Sigmodontinae) of South America. Oklahoma Museum of Natural History, Norman.
Czaplewski, N. J. 1987. Sigmodont rodents (Mammalia; Muroidea; Sigmodontinae) from the Pliocene (early Blancan) Verde Formation, Arizona. J. Vert. Paleont. 7:183-199.
Hershkovitz, P. 1959. Nomenclature and taxonomy of the neotropical mammals described by Olfers, 1818. J. Mamm. 40:337-353.
Hershkovitz, P. 1962. Evolution of neotropical cricetine rodents (Muridae) with special reference to the phyllotine group. Fieldiana: Zool. 46:1-524.
Mann, G. F. 1978. Los pequeños de mamíferos de Chile (marsupiales, quirópteros, edentados y roedores). Guyana (Zoología) 40:1-342.
Mares, M. A. 1977. Water balance and other ecological observations on three species of Phyllotis in northwestern Argentina. J. Mamm. 58:514-520.
Myers, P. 1977. A new phyllotine rodent (genus Graomys) from Paraguay. Occas. Pap. Mus. Zool. Univ. Michigan 1-7.
Olds, N., and S. Anderson. 1989. A diagnosis of the tribe Phyllotini (Rodentia, Muridae). 55-74 in Advances in Neotropical mammalogy (K. H. Redford and J. F. Eisenberg eds.). Sandhill Crane Press, Gainsville, Florida.
Olds, N., S. Anderson, and T. L. Yates. 1987. Notes on Bolivian Mammals 3: a revised diagnosis of Andalgalomys (Rodentia, Muridae) and the description of a new subspecies. Amer. Mus. Novitates 2890:1-17.
Osgood, W. H. 1947. Cricetine rodents allied to Phyllotis. J. Mamm 28:165-174.
Pearson, O. P. 1958. A taxonomic revision of the rodent genus Phyllotis. Univ. Calif. Publ. Zool. 56:391-477.
Pearson, O. P., and J. L. Patton. 1976. Relationships among South American phyllotine rodents based on chromosome analysis. J. Mamm. 57:339-350.
Simonetti, J. Z., and A. O. Spotorno. 1980. Posicion taxonomica de Phyllotis micropus (Rodentia: Cricetidae). An. Mus. Hist. Nat. Valparaiso 13:285-297.
Spotorno, A. O. 1986. Systematics and evolutionary relationships of Andean phyllotine and akodontine rodents. Ph.D. dissertation, University of California, Berkeley.
Steppan, S. J. 1993. Phylogenetic relationships among the Phyllotini (Rodentia: Sigmodontinae) using morphological characters. J. Mamm. Evol. 1:187-213.
Steppan, S. J. 1995a. Phylogenetic relationships of the phyllotine rodents (Sigmodontinae) and the evolution of phenotypic patterns of covariation in Phyllotis. Ph.D. dissertation, University of Chicago.
Steppan, S. J. 1995b. Revision of the leaf-eared mice Phyllotini (Rodentia: Sigmodontinae) with a phylogenetic hypothesis for the Sigmodontinae. Fieldiana: Zool. 80:1-112.
Walker, L. I., and A. E. Spotorno. 1992. Tandem and centric fusions in the chromosomal evolution of the South American phyllotines of the genus Auliscomys (Rodentia, Cricetidae). Cytogenet. Cell Genet. 61:135-140.
Information on the Internet
Mammal Species of the World: a Taxonomic and Geographic Reference. Internet version.Title Illustrations

| Scientific Name | Phyllotis darwini |
|---|---|
| Copyright | © 1996 P. L. Meserve |

| Scientific Name | Chinchillula sahamae |
|---|---|
| Comments | Chinchilla mouse |
| Copyright | © 1996 J. Anderson |
About This Page
Scott J. Steppan

Florida State University, Tallahassee, Florida, USA
Correspondence regarding this page should be directed to Scott J. Steppan at
Page copyright © 1996 Scott J. Steppan
All Rights Reserved.
- First online 01 October 1996
Citing this page:
Steppan, Scott J. 1996. Phyllotini. Leaf-eared mice and their relatives. Version 01 October 1996 (under construction). http://tolweb.org/Phyllotini/16592/1996.10.01 in The Tree of Life Web Project, http://tolweb.org/