Temporal fenestration has long been used to classify amniotes (Osborn, 1903). Taxa such as Anapsida, Diapsida, Euryapsida, and Synapsida were named after their type of temporal fenestration. Temporal fenestra are large holes in the side of the skull. The function of these holes has long been debated (Case, 1924), but no consensus has been reached. Many believe that they allow muscles to expand and to lengthen. The resulting greater bulk of muscles results in a stronger jaw musculature, and the longer muscle fibers allow an increase in the gape (Pirlot, 1969).
An important distinction must be made between the type of fenestration found in certain taxa, and the taxa that were named after these types of fenestration. For instance, the anapsid condition is characterized by the lack of temporal fenestrae (Figure 1). As such, it is primitive for amniotes because all their close relatives lack temporal fenestrae. Anapsida (the taxon) includes turtles and their extinct relatives. While most members of this taxon (but not lanthanosuchids and some millerettids) have an anapsid skull, some extinct relatives of saurians (such as captorhinids and "protorothyridids") also had anapsid skulls.

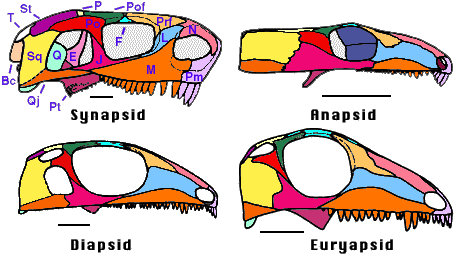
Figure 1. Amniote skulls in lateral view showing the four main types of fenestration. The synapsid condition is represented by the Lower Permian caseid Cotylorhynchus (Synapsida). The anapsid condition is represented by the Lower Periman amniote Captorhinus (Romeriida). The Pennsylvanian araeosceloid Petrolacosaurus (Diapsida) illustrates the diapsid condition, whereas its close relative, Araeoscelis (Diapsida) illustrates the euryapsid condition. Redrawn from the following sources: Synapsid, Laurin & Reisz (1995); Anapsid, Heaton (1979); Diapsid, Reisz, 1981; Euryapsid, Carroll, 1981. Abbreviations: Bc, braincase; E, epipterygoid; F, frontal; J, jugal; L, lacrimal; M, maxilla; N, nasal; P, parietal; Pm, premaxilla; Po, postrobital; Pof, postfrontal; Prf, prefrontal; Pt, pterygoid; Q, quadrate; Qj, quadratojugal; Sq, squamosal; St, supratemporal; T, tabular. Scale bars equal 1 cm.
The synapsid condition is characterized by the presence of a single temporal fenestra bordered minimally by the jugal, postorbital, and squamosal. The quadratojugal and the parietal occasionally contribute to the edge of this fenestra. By comparison with diapsids, this fenestra can be called a lower temporal fenestra. All early members of Synapsida, the taxon that includes mammals and their extinct relatives, had a synapsid skull, but the temporal fenestra has been drastically modified in mammals by ventral processes of the frontal and the parietal that occlude the temporal fenestra. The location of the old fenestra is still visible between the zygomatic arch, the orbit, and the dorsal part of the skull, but it is no longer a hole in the skull. A few taxa not related to Synapsida have also acquired a lower temporal fenestra. These include lanthanosuchids (Ivakhnenko, 1980) and some millerettids (members of Anapsida).
The diapsid condition is characterized by the presence of two temporal fenestrae, called the lower and the upper temporal fenestrae. The lower temporal fenestra is similar to the fenestra of synapsids, and it is generally bordered by the same bones (the jugal, postorbital, squamosal, and occasionally, the quadratojugal). The upper temporal fenestra is bordered by the postorbital, squamosal, parietal, and often, the postfrontal. This type of fenestration apparently appeared only once, in Diapsida, but it has been altered in many members of this taxon (squamates, and some early archosauromorphs, for example) by the loss of the lower temporal bar formed by the jugal and the quadratojugal.
The last type of fenestration called euryapsid has been the most problematic, partly because the origin of this condition has long been debated. It now appears that this condition is modified from the diapsid condition, and that it appeared more than once. Euryapsid skulls have only an upper temporal fenestra, usually bordered by the parietal, postfrontal, postorbital, and squamosal. Examples of euryapsid skulls include Araeoscelis, a Lower Permian araeoscelidian, placodonts, nothosaurs, and plesiosaurs (marine reptiles of the Mesozoic). Euryapsida is a taxon that includes most known euryapsids, except for Araeoscelis (Rieppel, 1993). The condition found in ichthyosaurs is sometimes distinguished from the euryapsid condition because their temporal fenestra is only bordered by the parietal, postfrontal, and supratemporal (Pirlot, 1969). This condition has been called parapsid, but it only represents a minor variation on the euryapsid pattern.

