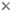Vampyroteuthidae
Vampyroteuthis infernalis
The Vampire Squid
Richard E. Young

Introduction
Vampyroteuthis infernalis is the only species known in the Vampyroteuthidae. The vampire squid is rather small, reaching a maximum of 21 cm ML (Hoving and Robison, 2012), and is very gelatinous; its consistency is that of a jellyfish. It occupies meso- to bathypelagic depths throughout the tropical and temperate regions of the world's oceans. The second pair of arms, apparently, is modified into retractile filaments that can extend to lengths up to eight times the total length of the animal, and they can be retracted into pockets between arms I and II (Hoving and Robison, 2012). The filaments appear to have a role in prey capture (Hunt, 1996), but in a novel fashion (see "Arms" next section). The vampire has black chromatophores with reddish-brown ones interspersed. [in most regions of the world's oceans, the black chromatophores dominate the general pigmentation, but off central California (e.g., the next two photographs) the reddish-brown cells dominate.] These chromatophores, however, have lost the muscles that enable the rapid color change seen in the chromatophores of other coleoids. A few normal coleoid chromatophores (i.e., chromatophore organs), however, are present on the vampire squid in the large, posterior photophores.
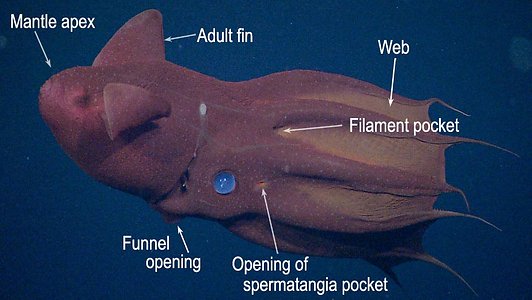
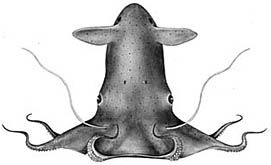
Figure. Two views of the same vampire squid showing some of their basic external features.© MBARI
The vampire sqid is a phylogenetic relict and possesses features of both octopodiforms and decapodiformes (most phylogenetic studies place it as an early branch of the Octopodiformes). In addition, it has many features that are probably adaptations to the deep-sea environment. Among these latter are the loss of the ink sac and most chromatophores organs, the development of photophores and the gelatinous consistency of the tissues.
Brief diagnosis:
An octopodiform ...
- with long, slender filaments (probably modified arms II).
- with cirri but without suckers on the proximal halves of arms.
Characteristics
- Arms
- Arms II (probably) modified to form retractile filaments (see details here).
- Lateral cirri present over arm length; suckers present only on distal half of arms where they alternate with cirri.
- Suckers without cuticular lining; sucker stalks constricted distally to narrow plate at base of sucker.
Click on an image to view larger version & data in a new window
Figure. Ventro-oral view of V. infernalis with arms folded aborally, showing cirri without associated suckers in proximal half of arms. Note the predominance of black pigment, compared to the previous two photographs, and the colorless, oral tips of the arms. Photographed in a shipboard aquarium in the North Atlantic by David Shale.
- Head
- Eyelid circular, without anterior optic sinus.
- Beaks: Descriptions can be found here: Lower beak; upper beak.
- Nuchal cartilage present even though head and mantle broadly fused.
- Central nervous system with incipient inferior frontal lobe system; superior buccal lobes adjacent (fused at edges) to posterior buccal lobes; suprabrachial commissure lies within brain.
- Eyelid circular, without anterior optic sinus.
- Funnel
- Funnel valve absent.
- Funnel valve absent.
- Mantle
- Dorsal mantle cavity absent.
- Mantle apex a highly-flexible muscular cone filled with gelatinous tissue located posterior to the cone of the gladius.
- Fins
- Two pairs of fins present during early ontogeny.
- Two pairs of fins present during early ontogeny.
- Spermatangia
- Receptacle (deep sac) for spermatangia located anterior to each eye in females.
- Receptacle (deep sac) for spermatangia located anterior to each eye in females.
- Shell
- Shell a gladius with broad median field and broad conus with or without a short, narrow cyclindrical rostrum.
- Shell a gladius with broad median field and broad conus with or without a short, narrow cyclindrical rostrum.
- Photophores
- Large circular, lidded organs (= "fin-base" organs or "large posterior photophores") present posterior to each adult fin .
- Numerous small organs distributed over ventral surfaces of mantle, funnel, head and aboral surface of arms and web (="skin-nodule" organs or "epidermal photophores"). Two patches on dorsal surface of head look like aggregrated small photophores (see images in Introduction) but are photoreceptors (Herring, et al., 1994) and may be "Nuchal Organs" (see Parry, 2000).
- Arm-tip organ(s) produce luminescent clouds consisting of microscopic glowing particles (Robison, et al., 2003).
- Arm-tip organs that flash or glow (Hunt, 1996).
- Pigmentation
- Viscera
- Oviducal glands at terminal end of oviducts.
- Visceropericardial coelom extends posteriorly as a slender duct, to a small organ at the apex of the conus of the gladius. This duct is possibly a remnant siphuncle.
- Photosensitive vesicles located immediately dorsal to funnel and, possibly, on dorsal integument in nuchal region.
Comments
A drawing and photographs of the mantle cavity can be seen here.
Nomenclature
A list of all nominal genera and species in the Vampyroteuthidae can be found here. The list includes the current status and type species of all genera, and the current status, type repository and type locality of all species and all pertinent references.
Life History
- Fins
- Development of the fins in the vampire is unique among cephalopods. One pair is present at hatching and, with only a slight increase in mantle length (if any), a second, more anterior fin-pair (the adult fins) develops giving the vampire squid four swimming fins. At about 25 mm ML the hatchling fins are quickly resorbed (Pickford, 1949). The first fin-pair to develop is the homologue of the fins of other cephalopods (Young and Vecchione, 1996). The unusual fin ontogeny is partially responsible for the early description of 3 families and many species where only one species actually exists. Except for the fins, the young vampire squid (about 10 mm ML) have an appearance very similar to that of the adult.
Click on an image to view larger version & data in a new window
Figure. The photograph shows an advanced four-fin stage in which the anterior, adult fins are much larger than the posterior hatchling-fins which will soon be resorbed. Note the mm scale, and the open large, posterior photophores. Photograph by John Bower.
- Eggs and hatchlings
- Vampires lack nidamental glands and have rather small oviducal glands. As a result there is little likelihood that they produce large egg masses. Off California small vampire squid occupy greater depths than do the larger individuals (Roper and Young, 1975) suggesting that spawning occurs in very deep water. A hatchling is known from deep water off Hawaii.
Behavior
Vampyroteuthis can swim surprisingly fast for a gelatinous animal. Hunt (1996) estimates from videotapes that it can reach two body lengths/sec, and it can accelerate to this speed in 5 sec. An escape reaction involves the quick movement of the fins toward the funnel followed by a jet from the mantle. This sequence is repeated as the vampire takes a series of quick turns in an erratic escape route (Hunt, 1996).




Figure. Vampyroteuthis infernalis. Sequence of images showing the fin movement during one fin stroke. The fin "stride" is finished with the fin pointing anteriorly and medially toward the funnel. The mantle intake for water at the lateral opening of the mantle cavity, is expanded in the first two images and closed in the last two, indicating that jet propulsion works in conjunction with the fin movement. Note the pointed posterior apex of the mantle seen during fast swimming. © MBARI. For more details on Vampyroteuthis swimming see: Seibel, 1998.
Postures
In slow swimming, the arms are sometimes spread anteriorly to form, along with the web, the shape of an umbrella or bell while the vampire slowly swims forward (Hunt, 1996). In another posture ("pineapple posture"), the arms and web are spread aborally over the head and mantle (Robison, 1995). In this defensive posture the squid would be somewhat more difficult to injure and would be covered by a densely pigmented cloak. The oral surface of the arms and webs are the most heavily pigmented (black) regions on the animal.
Bioluminescence
Figure. Vampyroteuthis infernalis filamentous, arm tip (preserved) showing the unpigmented, light-emitting surface. Note the loss ofl suckers and cirri on this arm tip (photograph by R. E. Young).
Hunt (1966) first observed bioluminescence displays in the living animal. The two large posterior photophores have been observed to glow brightly for less than a second (a flash) or longer than two minutes. The light intensity can vary giving a pulsating appearance, and, in addition, as light is extinguished, the glowing disc can be seen to decrease in diameter as well as intensity.
Arm-tips are unpigmented on their oral surface allowing light to be emitted from surfaces that have no obvious luminescent structures (see photograph on the right). These surfaces can glow, or they can flash at a rate of one to three per second or pulsate. With the arm-tip organs apparently glowing continuously, the vampire moves the arms ("arm writhing") around rapidly alternately exposing and hiding the luminescence which is "...very disorienting [to an observer] when trying to visually fix the animal's position" (Hunt, 1996 p. 104). Often a flash of the arm tips is followed by a rapid escape response. Another unusual and visually confusing effect is seen when viewing the vampire posteriorly (i.e., looking toward the mantle apex). The apparently disturbed vampire can curl the arms and web posteriorly over the head ("pineapple posture") then illuminate the arm-tip organs and the large posterior photophores. Hunt (1996, p. 104) states that the arm tips appear to come toward you, whereas the large posterior photophores appear to be moving away (due to their internal contraction) .
A third source of bioluminescence is the production of luminescent clouds whose function, seemingly, is to startle or distract a predator. These clouds appear as a mucous matrix with a few hundred to over 1000 discrete, glowing particles embedded in it. The particles can glow for up to 9.5 min. The source of the particles has been shown to be the arm tip organs (Robison, et al., 2003). The latter authors also suggest that flashing of the arm-tip organs is controlled by covering or exposing the photogenic region by the pigmented sides of the arm tips, and that the microscopic glowing particles of the luminescent clouds, are not luminescent bacteria.
Feeding
The vampire appears to orient most commonly in a horizontal attitude with generally one filament extended (Hunt, 1996). laboratory observations on living vampires suggest the filaments are tactile sense organs (Hunt, 1996) used in detecting prey. Hoving and Robison (2012) found that the filaments, also, aid in "capturing" prey which is primarily detritus.
Distribution
Vertical Distribution
Off California trawling data show most vampires between depths of 600-1100 m with peaks at 700-800 m and 900-1000 m, and with small individuals of less than 20 mm being most abundant at the deeper peak (Roper and Young, 1975). ROV observations in Monterey Bay, California suggest that the vampire occupies the oxygen minimum layer at depths of600-900 m with oxygen levels centered around 0.4 ml/l (Hoving and Robison, 2012).
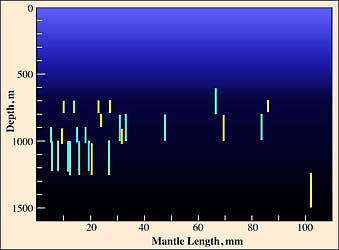
Vertical distribution chart modified from Clarke and Lu, 1975.
Off Hawaii, 10 of 11 captures came from depths of 800-1200 m but little towing was done in deeper water. Two captures were from opening-closing nets at depths of about 800-950 m.
In the Atlantic at 18° N, 25° W, the vampire shows a peak distribution between 700 and 1200 m but without a clear size/depth pattern (Clarke and Lu, 1975).
In the figure, all captures were made with opening/closing trawls. Bars represent a capture and the bar length indicates the depth range of the trawl while open. Yellow bars indicate a daytime capture and blue bars a nighttime capture. Fishing effort between 1000-1250 m was about twice that between 1250 and 1500 m, and effort between 1000-1500 m was about 5 times that between 1500 and 2000 m and about the same as that between 500 and 1000 m.
Numerous records exists for captures in excess of 1200 m (e. g., see Roper and Young, 1975) from open nets. Unfortunately, due to the rather high probability of contamination from shallower depths, these records are of questionable value.
Horizontal Distribution
The vampire squid is broadly distributed throughout the depths of the world's tropical and temperate oceans. Some geographical variation has been noted. Young (1972) found that the beaks of vampire squid from the Pacific Ocean off California were distinctly smaller than those from vampires of the Gulf of Guinea in the Atlantic Ocean. He also noted differences in sucker size and gill size in vampires from these areas. Vampires from off Monterey, California have a predominance of reddish rather than black chromatophores.
Other Names for Vampyroteuthis infernalis
- Vampyromorpha
- Vampyroteuthidae
- Vampyroteuthis
- The Vampire Squid
References
Clarke, M. R. and C. C. Lu. 1975. Verical Distribution of Cephalopods at 18° N 25° W in the North Atlantic. Journal of the Marine Biological Association of the United Kingdom, 55 (1): 165-182.
Herring, P. J., P. N. Dilly and C. Cope 1994. The bioluminescent organs of the deep-sea cephalopod Vampyroteuthis infernalis (Cephalopoda: Vampyromorpha). J. Zool. Lond. 233: 45-55.
Hunt, J. C. 1996. The behavior and ecology of midwater cephalopods from Monterey Bay: Submersible and laboratory observations. Ph. D. Dissertation, Univ. Calif. Los Angeles. 231 pp.
Hendrik J. T. Hoving and Bruce H. Robison. 2012. Vampire squid: detritivores in the oxygen minimum zone. Proc. R. Soc. B published online 26 September 2012 doi: 10.1098/rspb.2012.1357
Nesis, K. N. 1982. Abridged key to the cephalopod mollusks of the world's ocean. 385,ii pp. Light and Food Industry Publishing House, Moscow. (In Russian.). Translated into English by B. S. Levitov, ed. by L. A. Burgess (1987), Cephalopods of the world. T. F. H. Publications, Neptune City, NJ, 351pp.
Parry, M. 2000. A description of the nuchal organ, a possible photoreceptor, in Euprymna scolopes and other cephalopods. J. Zool. Lond. 252: 163-177.
Pickford, G. E. 1949. Vampyroteuthis infernalis Chun an archaic dibranchiate cephalopod. II. External anatomy. Dana-Report No. 32: 1-132.
Pickford, G. E. 1949. The distribution of the eggs of Vampyroteuthis infernalis Chun. Journal of Marine Research, 8(1):73-83.
Robison, B. H. 1995. Light in the ocean's midwaters. Scientific American 273:60-64.
Robison, B. H, K. R. Reisenbichler, J. C. Hunt and S. H. D. Haddock. 2003. Light production by the arm tips of the deep-sea cephalopod Vampyroteuthis infernalis. Biol. Bull., 205: 102-109.
Roper, C. F. E. and R. E. Young. 1975. Vertical distribution of pelagic cephalopods. Smithsonian Contributions to Zoolog, 209: 1-51.
Seibel, B. A., Thuesen, E. V. and Childress, J. J. 1998. Flight of the Vampire: Ontogenetic gait-transition in Vampyroteuthis infernalis (Cephalopoda: Vampyromorpha). J. exp. Biol. 201, 2413-2424.
Young, R. E. 1964. The anatomy of the vampire squid. Masters Thesis, Univ. of Southern Calif.
Young, R.E. 1967. Homology of retractile filaments of vampire squid. Science, 156(3782):1633-1634.
Young, R. E. 1972. The systematics and areal distribution of pelagic cephalopods from the seas off Southern California. Smithson. Contr. Zool., 97: 1-159.
Information on the Internet
Vampyroteuthis Videos
- Vampyroteuthis infernalis. National Geographic
- Deep Dark Ocean. Planet Earth
Title Illustrations

| Scientific Name | Vampyroteuthis infernalis |
|---|---|
| Location | San Clemente Basin off Southern California |
| Comments | Photographed in plankton kreisel aboard the R/V NEW HORIZON. Captured at 700 m depth. |
| Size | 25 cm total length |
| Copyright | © 1999 Brad Seibel |
| Scientific Name | Vampyroteuthis infernalis |
|---|---|
| Reference | from Young, R. E. 1972. The systematics and areal distribution of pelagic cephalopods from the seas off Southern California. Smithson. Contr. Zool., 97: 1-159. |
| Image Use |
 This media file is licensed under the Creative Commons Attribution-NonCommercial License - Version 3.0. This media file is licensed under the Creative Commons Attribution-NonCommercial License - Version 3.0.
|
| Copyright |
©
Richard E. Young

|
About This Page
Richard E. Young
University of Hawaii, Honolulu, HI, USA
Page copyright © 2019
 Page: Tree of Life
Vampyroteuthidae . Vampyroteuthis infernalis . The Vampire Squid.
Authored by
Richard E. Young.
The TEXT of this page is licensed under the
Creative Commons Attribution-NonCommercial License - Version 3.0. Note that images and other media
featured on this page are each governed by their own license, and they may or may not be available
for reuse. Click on an image or a media link to access the media data window, which provides the
relevant licensing information. For the general terms and conditions of ToL material reuse and
redistribution, please see the Tree of Life Copyright
Policies.
Page: Tree of Life
Vampyroteuthidae . Vampyroteuthis infernalis . The Vampire Squid.
Authored by
Richard E. Young.
The TEXT of this page is licensed under the
Creative Commons Attribution-NonCommercial License - Version 3.0. Note that images and other media
featured on this page are each governed by their own license, and they may or may not be available
for reuse. Click on an image or a media link to access the media data window, which provides the
relevant licensing information. For the general terms and conditions of ToL material reuse and
redistribution, please see the Tree of Life Copyright
Policies.
- Content changed 26 March 2019
Citing this page:
Young, Richard E. 2019. Vampyroteuthidae . Vampyroteuthis infernalis . The Vampire Squid. Version 26 March 2019. http://tolweb.org/Vampyroteuthis_infernalis/20084/2019.03.26 in The Tree of Life Web Project, http://tolweb.org/