



Osteoglossomorpha
Bonytongues
Guo-Qing Li and Mark V. H. Wilson


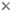
This tree diagram shows the relationships between several groups of organisms.
The root of the current tree connects the organisms featured in this tree to their containing group and the rest of the Tree of Life. The basal branching point in the tree represents the ancestor of the other groups in the tree. This ancestor diversified over time into several descendent subgroups, which are represented as internal nodes and terminal taxa to the right.

You can click on the root to travel down the Tree of Life all the way to the root of all Life, and you can click on the names of descendent subgroups to travel up the Tree of Life all the way to individual species.
For more information on ToL tree formatting, please see Interpreting the Tree or Classification. To learn more about phylogenetic trees, please visit our Phylogenetic Biology pages.
close boxIntroduction
The Osteoglossomorpha are a teleostean superorder defined by Greenwood, Rosen, Weitzman, and Myers in 1966. They are interesting to both ichthyologists and paleontologists because of their anatomy, physiology, geographic distribution, and ancient fossil record. Various osteoglossomorphs display unusual specializations of anatomy, such as elongate anal and dorsal fins and peculiarities of the jaws related to feeding, and of physiology and behaviour, such as generation of electric fields. Several species are also popular in the aquarium trade. Exclusively freshwater fishes at least in the modern fauna, they display interesting biogeographic distributions including apparent examples of endemism of extant suprageneric taxa (Hiodontidae in North America, Mormyroidea and Pantodontidae in Africa), as well as circumtropical or old-world tropical distributions (Osteoglossidae and Notopteroidea).
With regard to the fossil record, Eocene-age fossil members of genera now included in Osteoglossidae have been known for more than a century (Leidy 1873). Eocene fossil Hiodontidae have been recognized since Cavender’s (1966) work on †Eohiodon. However, the superorder was seen to be much older than Eocene when Greenwood (1970) suggested that the Asian fossil genus †Lycoptera, of Late Jurassic or Early Cretaceous age, might be an osteoglossomorph. Since 1970 numerous fossil genera from all major continents except Antarctica have been added to the list of potential extinct osteoglossomorphs. Examples include †Brychaetus Woodward (1901), †Paralycoptera Chang and Chou (1977), †Laeliichthys Santos (1985), †Chandlerichthys Grande (1986), †Yanbiania Li (1987), and †Ostariostoma Schaeffer (1949) as discussed by Grande and Cavender (1991) and Li and Wilson (1996a). Recent phylogenetic interpretations of these and other fossil taxa (Li 1994a-b, 1996, 1997; Li and Wilson 1994, 1996a-b, and in press; Li, Grande, and Wilson 1997; Li, Wilson, and Grande 1997) have added important information on the early divergence and the historical biogeography of the superorder.
Characteristics
Osteoglossomorph fishes share the following derived features:
- Primary bite between parasphenoid and tongue.
- Absence of supramaxillary bone.
- Absence of supraorbital bone.
- Fusion of fourth and fifth infraorbital bones.
- Number epurals bones decreased to one or zero.
- 18-17 or fewer principal caudal fin rays.
- Paired tendon bones on 2nd hypobranchial (in extant species).
- Intestine passes to the left of the stomach (in extant species).
Discussion of Phylogenetic Relationships
The Osteoglossomorpha consist of two orders containing living species, the Hiodontiformes or mooneyes and the Osteoglossiformes or bonytongues and relatives, along with two more primitive and exclusively fossil groups, the †Lycopteridae and †Kuyangichthidae. The two extant North American Hiodon species share two soft tissue character states with all other living osteoglossomorphs: 1) paired tendon bones on 2nd hypobranchial or 2nd hypobranchial and basibranchial, and 2) intestine coiling to the left of the esophagus and stomach (Nelson, 1972). These synapomorphies together define the monophyly of the Osteoglossomorpha (Li and Wilson, 1996b).
The order Hiodontiformes was erected by McAllister in 1968 and emended by Taverne in 1979. Since Bridge (1900) linked Hiodon with Notopterus based on the "swim bladder-ear connection", the Hiodontidae have been most often grouped in Notopteroidei with Notopteridae (Regan, 1909; Berg, 1940; Greenwood, 1963, 1970; Greenwood et al., 1966; Cavender, 1966; Patterson and Rosen, 1977; Grande, 1979; Lauder and Liem, 1983; J. S. Nelson, 1994), with the exception of Gosline (1960) and McAllister (1968). Recent study (see Li, 1994a; Li and Wilson, 1996a; Li, Wilson, and Grande, 1997) instead suggests a sister-group relationship between the Hiodontiformes and the Osteoglossiformes.
With the addition of the notopterids, the Osteoglossiformes form a well-defined monophyletic group supported by three synapomorphies:
- Nasal bone gutter-like or irregularly subrectangular
- Uroneural bones decreased to two or zero
- Branched caudal-fin rays 15 or fewer
The Asian fossil genus †Lycoptera Müller (1847) was formerly referred to the Esocidae or the †Leptolepidae (Woodward, 1901). Cockerell (1925) named the family †Lycopteridae and suggested that †Lycoptera might be “the ancestor of the Cyprinidae and their allies”. Berg (1940) grouped the family in Clupeiformes as suborder †Lycopteroidei; Yakovlev (1965) noted similarities between †Lycoptera and Arapaima, but Greenwood (1970) placed †Lycopteridae in the Hiodontoidea. Recent study suggests that †Lycopteridae are stem-group osteoglossomorphs, sister to all extant clades of the Osteoglossomorpha (Li and Wilson, 1996a, in press).
†Kuyangichthidae, named by Liu, Ma, and Liu (1982), probably consisting of †Tongxinichthys (Ma, 1980), †Kuyangichthys (Liu, Ma, and Liu , 1982), and †Jiuquanichthys (Ma, 1993), are tentatively considered to be a more primitive stem-group of Osteoglossomorpha, sister to †Lycopteridae plus all other osteoglossomorphs (Li and Wilson, in press). More evidence is needed concerning this possible relationship.
Relationship of Osteoglossomorpha to Other Teleostei
The systematic position of the Osteoglossomorpha has long been debated. Greenwood (1973) suggested a sister-group relationship between Osteoglossomorpha and Clupeomorpha, whereas Patterson and Rosen (1977), Lauder and Liem (1983), and J. S. Nelson (1994) considered Osteoglossomorpha to be the most primitive living teleosts. Arratia (1991) argued that elopomorphs are sister to osteoglossomorphs plus all other teleosts. We (Li and Wilson, 1996a) have not uncovered convincing synapomorphies for any of these alternatives; we therefore consider the extra-group relationships of the Osteoglossomorpha as still not well resolved.
References
Arratia, G. 1987. Anaethalion and similar teleosts (Actinopterygii, Pisces) from the Late Jurassic (Tithonian) of southern Germany and their relationships. Palaeontogra., (A), 200: 1-44.
Arratia, G. 1991. The caudal skeleton of Jurassic teleosts; a phylogenetic analysis. In "Early Vertebrates and Related Problems of Evolutionary Biology" (Chang M.-m., Liu Y.-h., and Zhang G.-r., eds.), pp. 249-340. Science Press, Beijing, China.
Arratia, G. 1996. Reassessment of the phylogenetic relationships of certain Jurassic teleosts and their implications on teleostean phylogeny. In “Mesozoic Fishes-Systematics and Paleoecology, Proceedings of International Meeting, Eichstätt 1993” (G. Arratia and G. Viohl, eds.), pp. 219-242. München (Verlag Dr. Pfeil).
Berg, L. S. 1940. Classification of Fishes both Recent and Fossil. Trudy Zool. Inst. Leningr., 5: 87-512 (English translation, Ann Arbor, Michigan, 1947).
Bridge, T. W. 1900. The air-bladder and its connection with the auditory organ in Notopterus borneensis. The Journal of the Linnean Society of London (Zoology), 27:503-540.
Cavender, T. (1966). Systematic position of the North American Eocene fish, "Leuciscus" rosei Hussakof. Copeia, 1966: 311-320.
Chang M.-m., and Chou C.-c. 1977. On Late Mesozoic fossil fishes from Zhejiang Province, China. Mem. Inst. Vert. Paleon. Paleoanthro., Acad. Sinica, 12: 1-59.
Cockerell, T. D. A. (1925). The affinities of the fish Lycoptera middendorffi. Bull. Amer. Mus. Nat. Hist., 51: 313-317.
Forey, P. L. 1973a. Relationships of elopomorphs. In "Interrelationships of Fishes" (P. H. Greenwood, R. S. Miles, and C. Patterson, eds.), pp. 351-368. Academic Press, London.
Forey, P. L. 1973b. A revision of the elopiform fishes, fossil and Recent. Bull. Brit. Mus. Nat. Hist. (Geol.), Supplement 10: 1-222.
Gosline, W. A. 1960. Contributions toward a classification of modern isospondylous fishes. Bulletin of the British Museum (Natural History), Zoology, 6:325-365.
Grande, L. 1979. Eohiodon falcatus, a new species of hiodontid (Pisces) from the late early Eocene Green River Formation of Wyoming. J. Paleont., 53: 103-111.
Grande, L. 1985. Recent and fossil clupeomorph fishes with materials for revision of the subgroups of clupeoids. Bull. Amer. Mus. Nat. Hist., 181: 231-372.
Grande, L. 1986. The first articulated freshwater teleost fish from the Cretaceous of North America. Palaeont. 29: 365-371.
Grande, L., and Cavender, T. M. 1991. Description and phylogenetic reassessment of the monotypic †Ostariostomidae (Teleostei). J. Vert. Paleont., 11: 405-416.
Greenwood, P. H. 1963. The swimbladder in African Notopteridae (Pisces) and its bearing on the taxonomy of the family. Bulletin of the British Museum (Natural History), Zoology, 11:377-412.
Greenwood, P. H. 1966. The caudal fin skeleton in osteoglossoid fishes. Ann. Mag. Nat. Hist., 9(13): 581-597.
Greenwood, P. H. 1970. On the genus Lycoptera and its relationship with the family Hiodontidae (Pisces, Osteoglossomorpha). Bull. Brit. Mus. Nat. Hist. (Zool.), 19: 257-285.
Greenwood, P. H. 1973. Interrelationships of osteoglossomorphs. In: "Interrelationships of Fishes" (P. H. Greenwood, R. S. Miles, and C. Patterson, eds.), pp. 307-332. Academic Press, London.
Greenwood, P. H. 1995. Bonytongues and their allies.In: "Encyclopedia of Fishes" (J. R. Paxton and W. N. Eschmeyer, eds.), pp. 80-84. Academic Press, San diego.
Greenwood, P. H., and Patterson, C. 1967. A fossil osteoglossoid fish from Tanzania (E. Africa). J. Linn. Soc. London (Zool.), 47: 211-223.
Greenwood, P. H., Rosen, D. E., Weitzman, S. H., and Myers, G. S. 1966. Phyletic studies of teleostean fishes, with a provisional classification of living forms. Bull. Amer. Mus. Nat. Hist., 131: 339-456.
Gregory, W. K. (1933). Fish Skulls: A Study of the Evolution of Natural Mechanisms. Trans. Amer. Philos. Soc., 23: 75-481.
Jin F. 1991. A new genus and species of Hiodontidae from Xintai, Shandong. Vert. PalAsi., 29: 46-54.
Kershaw, D. R. 1976. A structural and functional interpretation of the cranial anatomy in relation to the feeding of osteoglossoid fishes and a consideration of their phylogeny. Trans. Zool. Soc. London, 33: 173-252.
Lauder, G. V., and Liem, K. F. 1983. The evolution and interrelationships of the actinopterygian fishes. Bull. Mus. Com. Zool., 150: 95-197.
Leidy, J. 1873. Notice of remains of fishes in the Bridger Tertiary Formation of Wyoming. Proc. Acad. Nat. Sci. Philad., 25: 97-99.
LI, G.-Q. 1987. A new genus of Hiodontidae from Luozigou Basin, east Jilin. -Vert. PalAsit., 25: 91-107. (In Chinese with English abstract)
LI, G.-Q.1994a. New Osteoglossomorphs (Teleostei) from the Upper Cretaceous and Lower Tertiary of North America and Their Phylogenetic Significance. - Ph. D. Thesis, University of Alberta, Canada, 290 pp.
LI, G.-Q. 1994b. Systematic position of the Australian fossil osteoglossid fish †Phareodus (=†Phareoides) queenslandicus HILLS. -Mem. Queensl. Mus., 37: 287-300.
LI, G.-Q. 1996. A new species of Late Cretaceous osteoglossid (Teleostei) from the Oldman Formation of Alberta, Canada, and its phylogenetic relationships. In: ARRATIA, G. & VIOHL, G. (eds.): Mesozoic Fishes - Systematics and Paleoecology, p. 285-298. Verlag Dr. Pfeil, München.
LI, G.-Q. 1997. Notes on the historical biogeography of Osteoglossomorpha (Teleostei). In: Proceedings of the 30th International Geological Congress. VSP International Science Publishers, Zeist, The Netherlands: VoL. 12: 54-66.
LI, G.-Q. & WILSON, M. V. H. 1994. An Eocene species of Hiodon from Montana, its phylogenetic relationships, and the evolution of the postcranial skeleton in the Hiodontidae (Teleostei). -J. Vert. Paleont., 14: 153-167.
LI, G.-Q. & WILSON, M. V. H. 1996a. Phylogeny of Osteoglossomorpha. In: STIASSNY, M. L. J.; PARENTI, L. R.; & JOHNSON, G. D. (eds.): Interrelationships of Fishes, 1996: 163-174; San Diego, London, Boston, New York, Sydney, Tokyo, Toronto, 496 pp.
LI, G.-Q. & WILSON, M. V. H. 1996b. The discovery of Heterotidae (Teleostei: Osteoglossidae) from the Paleocene Paskapoo Formation of Alberta, Canada. - J. Vert. Paleont., 16: 198-209.
LI, G.-Q. & WILSON, M. V. H. (in press). Early divergence of Hiodontiformes sensu stricto in East Asia and phylogeny of some Late Jurassic -Early Cretaceous teleosts from China. In: ARRATIA, G. (ed.): Mesozoic Fishes - Systematics and the Fossil Records, p. 000-000. Verlag Dr. Pfeil, München.
LI, G.-Q.; GRANDE, L.; & WILSON, M. V. H. 1997. The species of †Phareodus (Teleostei: Osteoglossidae) from the Eocene of North America and their phylogenetic relationships. -J. Vert. Paleont., 17: 487-505.
LI, G.-Q.; WILSON, M. V. H. ; & GRANDE, L. 1997. Review of †Eohiodon (Teleostei: Osteoglossomorpha) from western North America, with a phylogenetic reassessment of Hiodontidae. J. Paleont., 71: 1109-1124. (USA)
Mcallister, D. E. 1968. Evolution of branchiostegals and classification of teleostome fishes. Bull. Nat. Mus. Can., 221: 1-239.
Maisey, J. G. 1991). Cladocyclus. In "Santana Fossils: An Illustrated Atlas" (J. G. Maisey, ed.), pp. 190-207. T. F. H. Publications, Inc., USA.
Müller, J. 1847. Fossile Fische. In "Reise in den aussersten norder und osten Sibiriens wahrend der Jahre 1843 und 1844 (Erster Band) (A. Th. Von Middendorff, ed.), pp. 260-264. Buchdruckerei der Kaiserlichen Akademie der Wissenschaften, St. Petersburg.
Nelson, G. J. 1968. Gill arches of teleostean fishes of the division Osteoglossomorpha. J. Linn. Soc. London (Zool.), 47: 261-177.
Nelson, G. J. 1969a. Gill arches and the phylogeny of fishes, with notes on the classification of vertebrates. Bull. Amer. Mus. Natu. Hist., 141: 475-552.
Nelson, G. J. 1969b. Infraorbital bones and their bearing on the phylogeny and geography of osteoglossomorph fishes. Amer. Mus. Nov., 2394: 1-37.
Nelson, G. J. 1972. Observations on the gut of the Osteoglossomorpha. Copeia, 1972: 325-329.
Nelson, J. S. 1994. Fishes of the World (3rd edition). John Wiley and Sons, New York. 600 pp.
Nybelin, O. 1974. A revision of the leptolepid fishes. Acta Reg. Soc. Sci. Litter. Goth., Zool.,
9: 1-202.
Patterson, C. 1967. Are the teleosts a polyphyletic group? Colloques Int. Cent. Nat. Rech. Scient., 163: 93-109.
Patterson, C., and Rosen, D. E. 1977. Review of ichthyodectiform and other Mesozoic teleost fishes and the theory and practice of classifying fossils. Bull. Amer. Mus. Nat. Hist., 158: 81-172.
Regan, C. T. 1909. The classification of teleostean fishes. Annals and Magazine of Natural History, Series 8, 3:75-86.
Ridewood, W. G. 1904. On the cranial osteology of the fishes of the families Mormyridae, Notopteridae, and Hyodontidae. J. Linn. Soc. London (Zool.), 29: 188-217.
Ridewood, W. G. 1905. On the cranial osteology of the fishes of the families Osteoglossidae, Pantodontidae, and Phractolaemidae. J. Linn. Soc. London (Zool.), 29: 252-282.
Santos, R. Da Silva. 1985. Laeliichthys ancestralis, novo gênero e espécie de Osteoglossiformes do Aptiano da Formaçao Areado, estado de Minas Gerais, Brasil. MME-DNPM, Geol. 27, Paleont. Estrat. 2: 161-167.
Schaeffer, Bobb. 1949. A teleost from the Livingston Formation of Montana. Amer. Mus. Nov., 1427: 1-16.
Shen, M. 1993. Fossil "osteoglossomorphs" in China and their implications on teleostean phylogeny. Abstracts, Symposium on Mesozoic fishes: Systematics and Palaeoecology, Jura-Museum, Eichstätt, Germany.
Taverne, L. 1979. Ostéologie, phylogénese et systématique des téléostéens fossiles et actuels du superordre des ostéoglossomorphes. Troisième Partie. Acad. Roy. Belg., Mém. Classe Sci., XLIII-Fascicule 3: 1-168.
Whitehouse, R. H. 1910. The caudal fin of the Teleostomi. Proc. Zool. Soc. Lond., 1910: 590-627.
Wilson, M. V. H. 1978. Eohiodon woodruffi n. sp. (Teleostei, Hiodontidae) from the middle Eocene Klondike Mountain Formation near Republic, Washington. Can. J. Earth Sci., 15: 679-686.
Wilson, M. V. H. 1992. Importance for phylogeny of single and multiple stem-group fossil species with examples from freshwater fishes. Syst. Biol., 41: 462-470.
Woodward, A. S. 1901. Catalogue of the Fossil Fishes in the British Museum (Natural History). Part IV. British Museum (Natural History), London. 636 pp.
Yakovlev, V. N. 1965. Systematics of the family Lycopteridae. Int. Geol. Rev., 1966, 8: 71-80 (English translation).
Title Illustrations

| Scientific Name | Osteoglossum, Phareodus |
|---|---|
| Location | South America (Osteoglossum), Green River Formation of Wyoming, USA (Phareodus) |
| Comments | Extant Osteoglossum from South America. Phareodus, the first named fossil osteoglossomorph from the Eocene Green River Formation of Wyoming, USA. |
| Specimen Condition | Fossil -- Period: Eocene (Phareodus) |
| Copyright | © 1998 Guo-Qing Li and Mark V. H. Wilson |
About This Page

Chinese Academy of Sciences, Beijing , China
University of Alberta, Edmonton, Alberta, Canada
Correspondence regarding this page should be directed to Guo-Qing Li at and Mark V. H. Wilson at
Page copyright © 1998 and
All Rights Reserved.
- First online 06 October 1998
Citing this page:
Li, Guo-Qing and Mark V. H. Wilson. 1998. Osteoglossomorpha. Bonytongues. Version 06 October 1998. http://tolweb.org/Osteoglossomorpha/15071/1998.10.06 in The Tree of Life Web Project, http://tolweb.org/