Hysteriales
Hysteriaceae
Eric Boehm







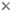
This tree diagram shows the relationships between several groups of organisms.
The root of the current tree connects the organisms featured in this tree to their containing group and the rest of the Tree of Life. The basal branching point in the tree represents the ancestor of the other groups in the tree. This ancestor diversified over time into several descendent subgroups, which are represented as internal nodes and terminal taxa to the right.

You can click on the root to travel down the Tree of Life all the way to the root of all Life, and you can click on the names of descendent subgroups to travel up the Tree of Life all the way to individual species.
For more information on ToL tree formatting, please see Interpreting the Tree or Classification. To learn more about phylogenetic trees, please visit our Phylogenetic Biology pages.
close boxIntroduction

Hysterothecium of Hysterium pulicare. ©
Fungi classified in the Hysteriaceae Chevall. (Chevallier 1826) are defined by a specialized ascocarp termed the hysterothecium (Clements 1909). Hysterothecia are dense, persistent carbonaceous structures, distinctly navicular in outline, and bear a pronounced longitudinal slit running the length of the long axis of the fruitbody. Hysterothecia may be capable of opening partially to reveal a lenticular, disk-like hymenium or closing tightly in response to relative humidity, suggesting that some may be perennial, capable of spore discharge over prolonged periods (Lohman 1933). They can be immersed to erumpent or entirely superficial, solitary or gregarious, ellipsoid to greatly elongated, and are sometimes branched, triradiate or borne on a subiculum (Zogg 1962). In vertical section, hysterothecia are globose to obovoid, typically with a thick three layered peridium, composed of small pseudoparenchymatous cells, the outer layer heavily encrusted with pigment and often longitudinally striated on the surface, the middle layer lighter in pigmentation and the inner layer distinctly thin-walled, pallid and compressed (Barr 1987). The hamathecium is composed of persistent cellular pseudoparaphyses, often borne in a gel matrix, with tips darkened or branched at maturity to form an epithecium. Bitunicate asci are borne in a basal layer and at maturity are typically clavate to cylindric, bearing eight ascospores, overlapping biseriate, ranging from hyaline to dark brown, obovoid, clavate, ellipsoid or fusoid. Ascospores are highly diverse in septation and range from didymospores to phragmospores to dictyospores, at times surrounded by a gel coating, and often show bipolar asymmetry (Barr 1987). The family has been monographed by Zogg (1962). Detailed taxonomic keys and illustrations can be found on the web at http://www.eboehm.com/.
Current classification of the Hysteriaceae (Eriksson 2006) includes the following genera: Hysterium Fr. : Fr., Hysterographium Corda, Gloniopsis De Not., Gloniella Sacc., Glonium Muhl. : Fr., Farlowiella Sacc., and Hysterocarina Zogg, to which has been recently added Actidiographium Vassiljeva. The genera Hysteroglonium Rehm ex Lindau, Hysteropatella Rehm and Pseudoscypha Reid & Piroz., are tentatively included in the Hysteriaceae by Eriksson (2006). In addition to these, the genera Encephalographa Massal., Graphyllium Clem., and Hemigrapha (Müll. Arg.) R. Sant. ex D. Hawksw. were included by Kirk et al. (2001). Both coelomycetous pynidial states (e.g., Hysteropycnis, Aposphaeria) and dematiaceous hyphomycetous anamorphs (e.g., Coniosporium, Septonema, Sirodesmium, Sphaeronaema and Sporidesmium) have been described (Lohman 1931, 1933, 1934). Hysteriaceous fungi are panglobal in distribution, having been reported from Japan (Amano 1983), China (Teng 1933), the Russian Far East (Vasilyeva 1999a, b, 2000), South Africa (Lee & Crous 2003; van der Linde 1992), Argentina (Messuti & Lorenzo 1997, 2003; Lorenzo & Messuti 1998), Costa Rica (Checa et al. 2007), North America (Lohman 1933, 1937) and Europe (Zogg 1962). Although primarily lignicolous or corticolous, recently a saxicolous / terricolous and apparently lichenized species has been described from Tasmania (Kantvilas & Coppins 1997).
Taxonomic History
The genus Hysterium, the type genus of the family Hysteriaceae, is attributed to Tode (1784), who was the first to apply the name to a group of fungi bearing a pronounced longitudinal slit, for which he gave the common name “Venusschwämme”. Recognizing the transitional nature of the ascoma, Tode later (1791) stated: “Medium hoc genus inter Pezizas & Lichenes”. Due to the seemingly transitional nature of the hysterothecium, neither fully open nor closed, hysteriaceous fungi have been placed in the discomycetes and pyrenomycetes about equally by various mycologists throughout the 19th Century (Bisby 1923). In his Systema Mycologicum, Fries (1823) initially considered hysteriaceous fungi to belong to the pyrenomycetes and placed them in the order Phacidiacei, but later (1835) placed them in his new class discomycetes, stating: “Transitum sistunt ad Discomycetes, sed discum verum non monstrant.” Ellis and Everhart (1892), in their North American Pyrenomycetes, tentatively included the Hysteriaceae, but stated that they had not at first intended to do so due to the transitional nature of the hysterothecium. Duby (1862) considered hysteriaceous fungi to belong to the pyrenomycetes and proposed two sections: the Hystériées to include Hysterium, Glonium, and Actidium Fr. among others, and the Lophiées to accommodate Ostreichnion Duby, Mytilinidion Duby and Lophium Fr. Although Duby’s (1862) method of classification, based on dehiscent versus nondehiscent asci, was not followed by subsequent workers, he was the first to propose dividing hysteriaceous fungi into what was later to become two distinct families. However, one hundred years would pass before this distinction was fully recognized (Zogg 1962).
Early authors did not follow Duby (1862) and instead classified fungi currently recognized as belonging to the Mytilinidiaceae within the family Hysteriaceae due to perceived similarities in ascocarp morphology, specifically its means of longitudinal dehiscence (Fries 1823; Ellis and Everhart 1892; De Notaris 1847; Massee 1895; Rehm 1896; Saccardo 1883; von Honel 1918). Modern authors have likewise included mytilinidiaceous fungi within the Hysteriaceae, placing the family in the Pseudosphaeriales (Nannfeldt 1932; Gäumann 1949), the Dothiorales (Müller & von Arx 1950; von Arx & Müller, 1954), the Dothideales (von Arx & Müller 1975) and in a separate order the Hysteriales, closely related to the Pleosporales (Miller 1949; Luttrell 1955). Luttrell (1953) studied ascomal ontogeny and hamathicial development in Glonium stellatum Mühlenb.:Fr. and concluded that the Hysteriaceae possess the pseudoparaphysate Pleospora-type centrum, in which cellular, septate pseudoparaphyses grow downwards from the cavity roof, initially anchored at both ends, and occupy the locule prior to the formation of asci (Luttrell 1951). The Hysteriales were placed in the subclass Loculoascomycetes by Luttrell (1955), due to the presence of bitunicate asci, corresponding to the Ascoloculares first proposed by Nannfeldt (1932). Zogg (1962) acknowledged the heterogeneity of the classical Hysteriales and, following Duby (1862), divided hysteriaceous fungi into two families, namely the Hysteriaceae to accommodate sessile, thick walled hysterothecia & the Mytilinidiaceae to accommodate modified hysterothecia, generally erect and possessing thin walls. Luttrell (1973) held a wide concept of the Hysteriales and did not recognize the family Lophiaceae, instead proposing a subfamily within the Hysteriaceae to accommodate mytilinidiaceous forms. Barr (1979) however maintained the two family distinction. The Mytilinidiaceae was placed in the Melanommatales, based on a thin-walled peridium of scleroparenchymatous cells enclosing a hamathecium of narrow trabeculate pseudoparaphyses, asci borne in a peripheral layer and with ascospores typically showing bipolar symmetry (Barr 1987, 1990). Barr (1983) eventually abandoned the Hysteriales and placed the Hysteriaceae within the Pleosporales due to the presence of cellular pseudoparaphyses, asci borne in a basal rather than peripheral layer and ascospores typically showing bipolar asymmetry. Kirk et al. (2001) maintained both the Hysteriaceae and the Mytilinidiaceae in the Hysteriales, but Eriksson (2006) removed the Mytilinidiaceae from the Hysteriales and considered it as Dothideomycetes et Chaetothyriomycetes incertae sedis, leaving the Hysteriaceae as the sole family in the Hysteriales.
Recently, Schoch et al. (2006), using a multigene phylogeny of the Dothideomycetes, based on the nuSSU and nuLSU, the transcription elongation factor (TEF1) and the RNA polymerase II second largest subunit (RPB2), provided evidence indicating that hysteriaceous fungi do not form a monophyletic group. Data from this preliminary analysis concluded that the Hysteriaceae should be regarded as Dothideomycetes incertae sedis. More recently, Boehm, Schoch & Spatafora (2008, unpublished), employing a much wider taxon sampling strategy, and using the same four genes, investigated whether morphological features historically used in the delineation of higher taxa in the Hysteriaceae were phylogenetically informative in the context of sequence-based phylogenies. It was found that the Hysteriaceae resides within the subclass Pleosporomycetidae, adjacent to the Pleosporales, and constitutes its own order, the Hysteriales.
References
Amano N, 1983. Saprobic loculoascoycetous fungi from Japan 1. Hysteriaceous fungi. Transactions of the Mycological Society of Japan 24: 283-297.
Arx JA von, Müller E, 1954. Die Gattungen der amerosporen Pyrenomyceten. Beiträge zur Kryptogamenflora der Schweiz 11(1): 1-434.
Arx JA von, Müller E, 1975. A re-evaluation of the bitunicate Ascomycetes with keys to families and genera. Studies in Mycology, Baarn 9: 1-159.
Barr ME, 1979. A classification of loculoascomycetes. Mycologia 71: 935-957.
Barr ME, 1983. The ascomycete connection. Mycologia 75: 1-13.
Barr ME, 1987. Prodromus to class Loculoascomycetes. Hamilton I. Newell, Inc., Amherst, Massachusetts: M.E. Barr Bigelow. 168 p.
Barr ME, 1990. Melanommatales (Loculoascomycetes). North American Flora, Series II, Part 13: 1-129.
Bisby GR, 1923. The literature on the classification of the Hysteriales. Transactions of the British Mycological Society 8: 176-189.
Checa J, Shoemaker RA, Umaña L, 2007. Some new hysteriaceous fungi from Costa Rica. Mycologia 99: 285-290.
Chevallier FF, 1826. Flore générale des environs de Paris, Vol I.
Clements FE, 1909. The Genera of Fungi.
De Notaris CG, 1847. Prime linee di una nuova disposizione de Pirenomiceti Isterini. Giornale Botanico Italiano 2, part I, fasc. 7-8: 5-52.
Duby JE, 1862. Mémoire sur la tribu des Hystérinées de la famille des Hypoxylées (Pyrénomycètes). Mémoires de la Société de Physique et Histoire Naturelle de Genève 16: 15-70.
Ellis JB, Everhart BM, 1892. The North American Pyrenomycetes. Newfield NJ. 793 p.
Eriksson OE. 2006. Outline of Ascomycota. Myconet 12: 1-88.
Fries EM, 1823. Systema Mycologicum, sistens fungorum ordines, genera et species hucusque cognitas, II, pars II: 276-620.
Fries EM, 1835. Corpus Florarum provincialium Sueciae, I. Floram Scanicam.
Gäumann EA, 1949. Die Pilze, Grundzüge ihrer Entwicklungsgeschichte und Morphologie. Birkhäuser. Basel, 382 p.
Höhnel F. von, 1918. Mycologische Fragmente, 272. Über die Hysteriaceen. Annales Mycologici 16: 145-154.
Kantvilas G, Coppins BJ, 1997. Melaspilea circumserpens Nyl. rediscovered and referred to Glonium, with discussion on the provenance of some of Robert Browns lichen specimens. Lichenologist 29: 525-531.
Kirk PM, Canon PF, David JC, Stalpers JA, 2001. Dictionary of the Fungi. 9th Ed. CAB International.
Lee S, Crous PW, 2003. Taxonomy and biodiversity of hysteriaceous ascomycetes in fynbos. South African Journal of Botany 69: 480-488.
Lohman ML, 1931. A study of Glonium parvulum in culture. Papers of the Michigan Academy of Science Arts & Letters 13: 141-156.
Lohman ML, 1933. Hysteriaceae: Life histories of certain species. Papers of the Michigan Academy of Science Arts & Letters 17: 229-288.
Lohman ML, 1934. A cultural and taxonomic study of Hysterium hyalinum. Papers of the Michigan Academy of Science Arts & Letters 19: 133-140.
Lohman ML, 1937. Studies in the genus Glonium as represented in the Southeast. Bulletin of the Torrey Botanical Club 64: 57-73.
Lorenzo LE, Messuti MI, 1998. Noteworthy Hysteriaceae from southern South America. Mycological Research 102: 1101-1107.
Luttrell ES, 1951. Taxonomy of the Pyrenomycetes. University of Missouri Studies in Science 24: 1-120.
Luttrell ES, 1953. Development of the ascocarp in Glonium stellatum. American Journal of Botany 40: 626-633.
Luttrell ES, 1955. The ascostromatic ascomycetes. Mycologia 47: 511-532.
Luttrell ES, 1973. Loculoascomycetes. In: The Fungi: an advanced treatise. IVA. Ainsworth GC, Sparrow FK, Sussman AS, eds. London, Academic Press.
Massee G, 1895. British Fungus Flora, Vol IV.
Messuti MI, Lorenzo LE, 1997. A new species of Hysterium from Patagonia, Argentina. Mycological Research 101: 302-304.
Messuti MI, Lorenzo LE, 2003. Notes on the genus Hysterographium (Ascomycota, Hysteriaceae) in southern South America. Nova Hedwigia 76: 451-458.
Miller JH, 1949. A revision of the classification of the Ascomycetes with special emphasis on the Pyrenomycetes. Mycologia 41: 99-127.
Müller E, von Arx JA, 1950. Einige Aspeckte zur Systematik pseudosphärialer Ascomyceten. Berichte der Schwizerischen Botanischen Gesellschaft 60: 329-397.
Nannfeldt JA, 1932. Studien über die Morphologie und Systematik der nicht-lichenisierten, inoperkulaten Discomyceten. Nova Acta Regiae Societatis Scientiarum Uppsaliensis IV, 8(2): 1-368.
Rehm H, 1896. Ascomyceten: Hysteriaceen und Discomyceten, In: L. Rabenhorsts Kryptogamen-Flora von Deutschland, Oesterreich und der Schweiz. 2nd Ed, Eduard Kummer, Leipzig 3: 1-56.
Saccardo PA, 1883. Sylloge Fungorum, omnium hucusque cognitorum 2: 776-783.
Schoch CL, Shoemaker RA, Seifert KA, Hambleton S, Spatafora JW, Crous PW, 2006. A multigene phylogeny of the Dothideomycetes using four nuclear loci. Mycologia 98: 1041-1052.
Teng SC, 1933. Notes on Hysteriales from China. Sinensia 4: 129-144.
Tode HJ, 1784. Beschreibung des Venusschwammes (Hysterium). Schriften der Berliner Gesellschaft Naturforschender Freunde 5: 53-55.
Tode HJ. 1791. Fungi Mecklenburgensis II.
van der Linde EJ, 1992. Notes on the South African Hysteriaceae (Ascomyctes: Mycotina). South African Journal of Botany 58: 491-499.
Vasilyeva LN, 1999a. Hysteriaceous fungi in the Russian Far East I. Hysterium. Mikologiya i Fitopatologiya 33: 225-227.
Vasilyeva LN, 1999b. Hysteriaceous fungi in the Russian Far East II. The genus Hysterographium. Mikologiya i Fitopatologiya 33: 297-301.
Vasilyeva LN, 2000. Hysteriaceous fungi in the Russian Far East III. Glonium and Actidiographium. Mikologiya i Fitopatologiya 34: 3-5.
Zogg H, 1962. Die Hysteriaceae s. str. und Lophiaceae, unter besonderer Berücksichtigung der mitteleuropäischen Formen. Beiträge zur Kryptogamenflora der Schweiz, Band 11, Heft 3: 1-190.
Title Illustrations
Gloniopsis praelonga (Schw.) Zogg

| Scientific Name | Gloniopsis praelonga (Schw.) Zogg |
|---|---|
| Location | Morris Co., NJ, USA, from Rubus. |
| Comments | Herbarium of EWA Boehm; Collected from dead Rubus canes, Morris Co, NJ, USA (Sept. 2002) |
| Specimen Condition | Dead Specimen |
| Identified By | EWA Boehm |
| Life Cycle Stage | Telomorph |
| Copyright |
©

|

| Scientific Name | Gloniopsis praelonga (Schw.) Zogg |
|---|---|
| Location | Morris Co., NY, Rubus, 2004. |
| Comments | Hyaline dictyospores define the genus Gloniopsis. |
| Specimen Condition | Live Specimen |
| Identified By | E.W.A. Boehm |
| Copyright |
©
|

| Scientific Name | Hysterographium flexuosum (Schw.) Sacc. |
|---|---|
| Location | Union Co., NJ, USA. 2007. |
| Comments | Collected from Quercus sp. Watchung Reservation, Union Co., NJ, USA. 2007. |
| Specimen Condition | Live Specimen |
| Identified By | E.W.A. Boehm |
| Copyright |
©
|

| Scientific Name | Hysterographium flexuosum (Schw.) Sacc. |
|---|---|
| Location | Union Co., NJ, USA. 2007. |
| Specimen Condition | Live Specimen |
| Identified By | E.W.A. Boehm |
| Copyright |
©
|

| Scientific Name | Hysterium sinense Teng |
|---|---|
| Location | Watchung Reservation, Union Co., NJ |
| Comments | This is the first report of Hysterium sinense from the New World, having been previously described from China and South Africa. |
| Identified By | Eric W.A. Boehm |
| Copyright |
©
|

| Scientific Name | Hysterium sinense Teng |
|---|---|
| Identified By | Eric W.A. Boehm |
| Copyright |
©
|
About This Page
Kean University, Union, New Jersey, USA
Correspondence regarding this page should be directed to Eric Boehm at
Page copyright © 2008
 Page: Tree of Life
Hysteriales. Hysteriaceae.
Authored by
Eric Boehm.
The TEXT of this page is licensed under the
Creative Commons Attribution-NonCommercial License - Version 3.0. Note that images and other media
featured on this page are each governed by their own license, and they may or may not be available
for reuse. Click on an image or a media link to access the media data window, which provides the
relevant licensing information. For the general terms and conditions of ToL material reuse and
redistribution, please see the Tree of Life Copyright
Policies.
Page: Tree of Life
Hysteriales. Hysteriaceae.
Authored by
Eric Boehm.
The TEXT of this page is licensed under the
Creative Commons Attribution-NonCommercial License - Version 3.0. Note that images and other media
featured on this page are each governed by their own license, and they may or may not be available
for reuse. Click on an image or a media link to access the media data window, which provides the
relevant licensing information. For the general terms and conditions of ToL material reuse and
redistribution, please see the Tree of Life Copyright
Policies.
- First online 09 January 2008
- Content changed 25 June 2008
Citing this page:
Boehm, Eric. 2008. Hysteriales. Hysteriaceae. Version 25 June 2008 (under construction). http://tolweb.org/Hysteriaceae/29322/2008.06.25 in The Tree of Life Web Project, http://tolweb.org/